Professional Documents
Culture Documents
Micagricap 1
Uploaded by
kab22Original Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Micagricap 1
Uploaded by
kab22Copyright:
Available Formats
Leonor Carrillo. 2003. Microbiologa Agrcola.
captulo 1
1. Microorganismos
Los ambientes capaces de albergar vida microbiana reflejan el amplio espectro de la evolucin de estos organismos. Se han encontrado especies que viven a temperaturas comprendidas entre el punto de congelacin y el punto de ebullicin del agua, en agua salada y en agua dulce, en presencia y en ausencia de aire. Algunos han desarrollado ciclos de vida que incluyen una fase de latencia en respuesta a la falta de nutrientes. Los microorganismos se hallan capacitados para acometer una extensa gama de reacciones metablicas y adaptarse a muchos ambientes diferentes (1). Por su poco peso pueden ser transportados por las corrientes de aire y estar en todas partes, pero las caractersticas del ambiente determinan cules especies pueden multiplicarse. Existen varias clases de microorganismos: mohos, levaduras, bacterias, actinomicetos, protozoos, algas, virus. La figura 1 muestra el tamao comparativo de algunos microorganismos. El suelo es uno de los ambientes donde un conjunto ingobernable de microorganismos compiten entre s para obtener lo que todos ellos necesitan: nutrientes y energa. Al mismo tiempo, los productos de su metabolismo alteran la composicin qumica del suelo donde habitan. Ms an, los propios microorganismos evolucionan en respuesta a la presin del ambiente (3). En un suelo agrcola estn presentes alrededor de 1010 organismos por g de suelo y constituyen una biomasa de aproximadamente 1500 kg por Ha, lo cual corresponde a un cordero por cien m2 . Un gramo de suelo frtil puede contener 5 m de micelio fngico, 10 8 clulas bacterianas, 106 esporos de actinomicetos (4). En los animales monogstricos la poblacin bacteriana alcanza su mximo nivel en el intestino grueso y el impacto metablico de la microbiota es considerable. En el rumiante la accin de los microorganismos contenidos en el rumen es an ms espectacular, pues al degradar la celulosa permiten al animal alimentarse de forrajes (5). Numerosas especies bacterianas y algunas veces incluso algas microscpicas o protozoos, proliferan en las superficies expuestas a la humedad formando una biopelcula de microorganismos contenidos en una matriz de polisacridos, cuyo espesor puede oscilar entre algunos micrmetros y pocos milmetros, que se adhiere fuertemente a la base. Las biopelculas se forman en todas las superficies sumergidas, tanto en agua dulce como de mar, o bien sobre soportes constantemente hmedos tales como paredes de la caera de agua, pisos o dientes. El 99% de toda la actividad microbiana en un ecosistema abierto ocurre sobre las superficies (6). Los mohos y levaduras forman parte de los hongos. Las levaduras son unicelulares en condiciones normales mientras que los mohos crecen como un sistema ramificado de filamentos (micelio). Se trata de organismos eucariticos cuyas clulas, al igual que las
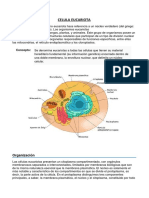
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1 de vegetales y animales, tienen un ncleo cerrado por una doble membrana porosa y llevan como mnimo dos cromosomas, a ms de veinte en algunas especies (7). Las eubacterias y los actinomicetos son procariticos. Sus clulas carecen de membrana nuclear y mitocondrias, adems poseen un solo cromosoma que se encuentra libre en el citoplasma y una pared celular rgida. Las clulas de los procariotas son, en general, mucho menores que las de los eucariotas; unas 10 veces menor en medidas lineales y por lo tanto unas 1000 veces menor en volumen (1). Los protozoos del suelo se alimentan de bacterias y presentan movimientos ligados a la fagocitosis que es la ingestin de partculas mediante invaginacin de la membrana citoplasmtica y formacin de vesculas intracelulares (4). Las eubacterias presentan diversa morfologa: esfrica, cilndrica, espiralada e incluso filamentosa. Los actinomicetos incluyen muchos gneros que desarrollan en forma de delgado micelio. Las eubacterias son organismos pequeos, que miden uno o varios micrmetros. Las levaduras, en cambio, miden entre 6 y 12 m. Los mohos son pluricelulares y aunque sus clulas aisladas miden a lo sumo 25 m de largo, su conjunto se distingue a simple vista. Los hongos con reproduccin sexual pueden tener cuerpos fructferos de tamao notable como los championes y otras setas, formados por millones de clulas (7). La figura 2 es un esquema de las clulas procaritica y eucaritica.
1.1. Dominios y reinos
Las dificultades lgicas para ubicar a los microorganismos en los vegetales o los animales desaparecieron al admitir un tercer reino, el de los protistas. Este reino inclua a todos aquellos organismos que se diferencian de las plantas y los animales, por su falta de especializacin morfolgica, siendo la mayora unicelulares. Luego los protistas fueron divididos en dos grupos claramente definidos sobre la base de su estructura celular. Los eucariticos eran protistas superiores. Este grupo comprenda a los protozoos, hongos y algas. Los procariticos eran protistas inferiores e incluan a las diversas bacterias y cianobacterias (2). Otra clasificacin de los seres vivos estaba fundada en el reconocimiento de cinco reinos (9).
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
Los estudios ultraestructurales, bioqumicos y de la biologa molecular hicieron insostenible la permanencia de estos reinos, y los procariotas fueron ubicados en los dominios Bacteria y Archaea mientras que el dominio Eukarya comprende a los microorganismos en tres reinos (10). La importancia de esto ltimo radica en que los organismos estudiados por los fitopatlogos han sido distribuidos entre los reinos Chromista, Fungi y Protozoa, aunque se ha sugerido transferir los hongos que estn en Chromista al nuevo reino Straminipila (11). Los virus no fueron considerados pues son partculas acelulares, diferentes de todos los otros microorganismos, que pueden proliferar y replicarse solamente dentro de clulas vivas (12). Por otra parte, parece que las clulas eucariticas surgieron de antepasados procariticos, pues hay pruebas del origen bacteriano de mitocondrias y cloroplastos (13). Dominio Archea Bacteria Eukarya Reino Tipo celular procaritico " eucaritico " " " " Ejemplos arquibacterias eubacterias, actinomicetos, mixobacterias, cianobacterias oomicetos, algas mixomicetos, protozoos levaduras, mohos, setas, bejines musgos, hepticas, conferas, plantas con flores esponjas, corales, gusanos, moluscos, insectos, vertebrados
Chromista Protozoa Fungi Plantae Animalia
1.2. Organismos procariticos
La clula procaritica tiene solamente dos sistemas internos principales: una molcula circular de ADN con cadena helicoidal doble y un citoplasma no diferenciado donde se halla inmerso ese ADN. La longitud del anillo de ADN que codifica toda la informacin gentica de la clula apenas es superior a un milmetro (7). El citoplasma contiene gran nmero de ribosomas, grnulos constituidos por ARN y protenas, que cumplen la funcin de enlazar los aminocidos formando protenas. Los ribosomas procariticos son ms pequeos que los eucariticos (1). La replicacin del ADN comienza cuando ambas cadenas se separan, permitiendo la sntesis de una cadena complementaria a cada una de ellas mediante la ADN polimerasa. La nueva hlice es un hbrido consistente de una cadena original y una recin formada (replicacin conservativa) (14). La duplicacin del cromosoma de Escherichia coli en
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1 condiciones favorables tarda 20 minutos. Un gen, o unidad de informacin gentica, est representado por una secuencia especfica del ADN y determina la estructura de un polipptido. El conjunto de genes se denomina genoma. La informacin contenida en la secuencia del ADN es transferida al ARN mensajero (ARN-m) durante la transcripcin. Los aminocidos son reunidos en una cadena polipeptdica segn la secuencia determinada por el ARN-m en un proceso de traduccin que implica la participacin del ARN-t (ARN que transfiere los aminocidos), los ribosomas, varias enzimas y ATP. Los microorganismos procariticos poseen una cubierta celular muy diferente de la de los eucariotas. Las paredes celulares de muchos de los procariotas poseen un componente qumico comn, el pptidoglucano (murena), que es el responsable de la forma y consistencia de la pared. El pptidoglucano es un extenso polmero que se halla integrado por subunidades alternas de N-acetilglucosamina (que es tambin el constituyente de la quitina en los eucariotas) y el cido N-acetilmurmico. Esta ltima molcula es similar a la N-acetilglucosamina, pero tiene una unidad de cido lctico unida al tercer tomo de carbono de la glucosa. El cido lctico sirve como punto de unin de una cadena lateral lineal constituda por cuatro aminocidos D o L. Las molculas de pptidoglucano que rodean la clula estn entrelazadas por puentes formados por los aminocidos (1). Algunas bacterias filamentosas forman una envoltura tubular constituda por heteropolisacridos que se conoce como vaina (12). La flexibilidad metablica de una bacteria es enorme. Debido al pequeo tamao, una bacteria esfrica tiene espacio para no ms que cien mil molculas de protenas. Las enzimas no estn corrientemente dentro de la clula, sino que su sntesis es inducida por la presencia del substrato en el ambiente (1).
1.2.1. Eubacterias
La cantidad de pptidoglucano de la pared de las clulas bacterianas vara enormemente, desde el 90% en la pared de algunas bacterias gram-positivas hasta menos del 10% en las bacterias gram-negativas. El trmino Gram-positivo hace referencia a la capacidad de ciertas bacterias de retener el colorante violeta cristal en presencia de yodo cuando se agrega alcohol, al hacer la coloracin de Gram sobre un portaobjetos con las bacterias fijadas. Las bacterias negativas no retienen a este colorante (7). La penicilina y otros antibiticos -lactmicos que interfieren con la biosntesis de la pared celular llevan a la produccin de protoplastos carentes de pared bajo condiciones osmticas apropiadas (1). El trmino esferoplasto se usa para los casos en que persisten restos de pared. La figura 3 muestra un esquema de las paredes celulares. La pared celular de las bacterias Grampositivas contiene pequeas cantidades de protenas y polisacridos, a menudo tambin cidos teicoicos (polmeros de ribitol-fosfato o glicerol-fosfato unidos mediante enlaces fosfodister) o de cidos teicournicos. Estos cidos estn unidos al pptido-glucano por los fosfatos (12). En las proteobacterias (Gramnegativas) la capa interna de la pared celular es pobre en pptidoglucano y la capa externa rica en lipoprotena y lipopolisacridos, los cuales constituyen ms del 80% del peso seco de la pared. El espacio entre la membrana externa y la capa de pptidoglucano, llamado periplsmico, contiene un gran nmero de protenas
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
enzimticas. La capa de pptido-glucano e st rodeada por una membrana externa compuesta de lpido A y fosfolpidos con protenas en tnel (porinas) que la atraviesan. El lpido A consiste de un disacrido de glucosamina cuyos grupos hidroxilo estn esterificados con cidos grasos de 12, 14 16 carbonos (12). Los micoplasmas son bacterias carentes de pared celular y parecen protoplastos, pero son ms resistentes a la lisis osmtica debido a la naturaleza de su membrana citoplasmtica, la que contiene esteroles en un grupo y carotenoides u otros compuestos en los dems (1). La membrana citoplasmtica bacteriana contiene alrededor del 70-90% de los lpidos celulares mientras que la membrana constituye solamente 8-15% del peso celular seco. La membrana, con un grosor de 5 nanmetros, consiste de una doble capa lipdica con los extremos hidrofbicos de los fosfolpidos en el interior y las cabezas hidroflicas expuestas sobre ambas superficies. Tambin tiene incorporadas protenas que la atraviesan o estn inmersas parcialmente en ella. Otras protenas unidas a la membrana son conocidas como protenas perifricas (17). Las substancias hidrofbicas difunden fcilmente pero no los iones. Las protenas de la membrana intervienen en la entrada y salida de substancias de la clula, y son especficas para un grupo de molculas estrechamente relacionadas. En la figura 4 se observa un esquema de la membrana citoplasmtica La membrana citoplasmtica se extiende y repliega hacia el interior del citoplasma formando los sitios de la generacin de energa respiratoria (mesosomas) y fotosinttica (tilacoides y cromatforos) en las bacterias que posean tales actividades metablicas. Otras bacterias (por ejemplo Nitrobacter, Nitrosomonas, Nitrosococcus) tienen paquetes de lminas paralelas, algunas de las cuales estn conectadas a la membrana citoplasmtica (12). Muchas bacterias estn dotadas de flagelos que les permiten moverse, insertados en la superficie celular. Si los flagelos se encuentran concentrados en un extremo de la clula se denominan polares ; si estn distribudos por toda la superficie celular, reciben el nombre de peritricos (7). La figura 5 muestra la ubicacin de los flagelos en las bacterias. Los flagelos posibilitan la quimiotaxis, que es el movimiento hacia la fuente de nutrientes o el alejamiento de un entorno qumicamente hostil. La protena del flagelo, flagelina, est unida a un gancho asociado a un cuerpo basal que causa el movimiento giratorio del flagelo. La base del flagelo est anclada en la membrana citoplasmtica, asociada en las gram negativas al pptido glucano y a la membrana externa para estabilizar la estructura flagelar (1). Las bacterias sin flagelos carecen de movimiento excepto aqullas que se deslizan sobre el substrato. La superficie bacteriana gram negativa tambin suele tener otras estructuras proteicas filamentosas las fimbrias o pili I que participan en la adhesin a diversas superficies, por ej. las mucosas animales, y los pelos de conjugacin o pili F (18). A veces las bacterias acumulan capas de poliscaridos sobre la superficie exterior (ej. Xanthomonas), pero algunas tienen una cpsula polipeptdica (ej. Bacillus). La cpsula no es esencial para la vida y no siempre est presente. Para observarla se suspende al microorganismos en una solucin de nigrosina o rojo Congo y como la cpsula no es teida se ve como un halo claro alredor de la clula. En muchos casos el material capsular puede ser separado en forma de limo, por agitacin u homogeinizacin. Por ejemplo, Leuconostoc mesenteroides convierte rpidamente a una solucin de azcar de caa en una jalea de dextrano (1,6--glucano). Por otra parte Acetobacter aceti var. xylinum secreta celulosa formado una cubierta coricea que rodea a las clulas y da cohesin a la colonia (12).
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1 Numerosas eubacterias y tambin arqueobacterias, poseen una envoltura exterior proteica o capa S que suelen perder en las condiciones ptimas del laboratorio. Esta capa favorece la adhesin al substrato para permitir la accin de las exoenzimas y a la virulencia de las bacterias patgenas (19). Un pequeo grupo de eubacterias que se encuentran en el suelo (Bacillus, Clostridium, Sporosarcina, etc.) poseen endosporos. Son paquetes de ADN y otras molculas que pueden permanecer en estado latente por muchos aos, y resultan altamente resistentes al calor (incluso al agua hirviendo), productos quimiotxicos y otros agentes que ocasionan la muerte a las clulas. Ya en el ambiente apropiado, el endosporo puede dar origen a una nueva bacteria (7). Si se calienta una suspensin de suelo a 80-100C durante 10 minutos, slo sobreviven los endosporos bacterianos. stos contienen cido dipicolnico, una substancia no hallada en la forma somtica, en la proporcin de 10-15% del peso seco del esporo. Son varias las cubiertas que rodean al citoplasma del endosporo: membrana citoplasmtica, pared celular, crtex (pptidoglucanos), envoltura interna, envoltura externa (polipptidos) y, en ocasiones, exosporio (10). La figura 6 muestra un esquema de la formacin y estructura de un endosporo bacteriano. Los plsmidos son pequeos anillos (hay algunos lineales) de ADN extracromosomal que se multiplican independientemente en las clulas que los alojan y pueden portar genes que confieren ciertas ventajas a las mismas, por ejemplo resistencia a los antibiticos. Las bacterias hospedadoras a su vez, dejan que el ADN plasmdico se replique en ellas hasta cierto punto, asegurando as la presencia continuada del plsmido en las clulas hijas (20). Los tumores de las plantas producidos por la infeccin con Agrobacterium tumefaciens se deben al plsmido Ti que es el agente infeccioso real. El ADN plasmdico penetra en la clula vegetal y parte del mismo se incorpora al genoma de la planta. Este proceso se llama transformacin oncognica. La parte integrada del plsmido lleva la informacin para la produccin de opinas que son substancias utilizadas solamente por la bacteria. Otra especie, Agrobacterium rhizogenes desencadena el crecimiento anormal de las races mediante el plsmido Ri (21). La conjugacin es una forma de transferencia de genes y para que se produzca debe haber contacto entre las clulas bacterianas a travs de los pili F o pelos de conjugacin que retienen juntas a las clulas dadora y receptora. Los sistemas de conjugacin en bacterias gram-negativas produce la transferencia de una parte del ADN del cromosoma o de un plsmido de la clula dadora a la receptora (20). Las bacterias forman colonias caractersticas sobre la superfice del substrato, que tienden a adoptar una forma circular creciendo por adicin de clulas a su permetro. Al extenderse la colonia sobre el agar se observa que en el crecimiento participan unos
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
elementos en anillos y otros en sectores. A todos los sectores los va formando, por propagacin centrfuga, la progenie de un antepasado comn. Algunos sectores se diferencian del resto porque las clulas difieren en su ADN de las de los sectores contiguos debido a mutaciones espontneas. Las bacterias de un anillo comparten algunas propiedades entre s pero no estn emparentadas, pues tienen parentesco directo con las de la zonas precedente y la siguiente (9). La figura 7 presenta algunas colonias de bacterias de distinto aspecto.
Las bacterias generalmente se multiplican por fisin binaria. Despus del crecimiento de la clula, aparece el septo y las clulas se separan o permanecen unidas con formas caractersticas: pares de bacterias, cadenas, paquetes cbicos y acmulos planos irregulares entre las bacterias esfricas (cocos), o pares y cadenas en las cilndricas (bacilos) (12). La multiplicacin por brotacin es rara en los procariotas pero se observa, por ejemplo, en la bacteria del agua estancada Hyphomicrobium. La figura 8 muestra un esquema de las diversas formas de bacterias. La segregacin del ADN es el proceso que distribuye igualitariamente el material gentico en las clulas hijas. Es producida en las bacterias debido a un mecanismo parecido al mittico. La replicacin de los plsmidos en la clula, sobre el replisoma, entraa la formacin de un complejo de separacin o particin donde se adhiere el plsmido cual regin centromrica. Una vez formadas las copias, stas se alejan activamente unidas a las protenas de particin hasta que la clula se divide. La divisin del cromosoma parece ocurrir de igual manera (23). La figura 9 presenta un esquema de la divisin de una bacteria.
1.2.2. Arqueobacterias
Este grupo de procariotas en su bioqumica y en la estructura de los ribosomas, membrana y pared, difieren tanto de los procariotas como de los eucariotas. Se los denomina arqueobacterias pues algunas poseen un metabolismo particularmente adecuado a las condiciones que se supone prevalecieron en los primeros tiempos de la vida sobre la tierra. Comprende tres clases muy diferentes: las metangenas, las halfilas extremas y las termoacidfilas (8). Las arqueobacterias no tienen pptidoglucano en la pared. La membrana est formada por lpidos no habituales, compuestos por un grupo glicerol ligado a dos cadenas de alquil-isoprenoides por un enlace tipo ter ( -O-) (24). Algunas de estas bacterias tiene histonas y nucleosoma de tipo eucaritico (25).
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1 Las metangenas ( ethanobacterium, etc.) viven solamente en ambientes libres de M oxgeno y liberan metano mediante la reduccin del CO2 . Su apariencia externa es la de las dems bacterias pero su pared celular vara. Algunas metanobacterias contienen pseudomurena donde el cido acetilmurmico est reemplazado por el cido Nacetilalosaminurnico pero en otras predominan las protenas o los heteropolisacridos en la pared celular. Viven en estrecha asociacin con otras bacterias, como los clostridios, que metabolizan materia orgnica en descomposicin y desprenden hidrgeno como producto de desecho. Se encuentran en agua estancadas, en las plantas de tratamiento de aguas residuales, en la panza del ganado, en el intestino de los animales, en el fondo del ocano y en los manantiales de aguas termales (1) (ver captulo 5). Las halfilas extremas son bacterias que, para sobrevivir, requieren elevadas concentraciones de sal. Algunas de ellas crecen fcilmente en salmuera saturada. Pueden conferir color rojo a los estanques de evaporacin donde se obtiene la sal y alteran al pescado salado. Mantienen fuertes gradientes en la concentracin de ciertos iones a travs de la membrana celular y utilizan esos gradientes para transportar diversas substancias hacia adentro o hacia afuera de la clula. Poseen un mecanismo fotosinttico sencillo que se basa en un pigmento ligado a la membrana: la rodopsina bacteriana. La pared de Halobacterium est formada por protenas pero la de Halococcus por heteropolisacridos (12). Entre las termoacidfilas se encuentra Sulfolobus que se halla en los manantiales de aguas termales sulfurosas. Crecen a ms de 80C y a pH inferior a 2. Otro gnero es Termoplasma, un micoplasma que slo posee membrana celular y crece a ms de 55C. El medio interior de la clula tiene un pH prximo a la neutralidad; ello exige mantener un considerable gradiente de pH a travs de la membrana celular. El gradiente es empleado para bombear molculas hacia dentro o fuera de la clula (8).
1.2.3 . Actinomicetos
Los actinomicetos constituyen un importante grupo de organismos procariticos habitantes del suelo y del material vegetal compostado. El gnero principal del grupo es Streptomyces cuyas especies suelen excretar antibiticos y el olor a tierra mojada se debe a compuestos voltiles fabricados por los mismos. Cuando se los cultiva en medio slido, no slo forman un fino micelio ramificado, sino que tambin producen una hifa area que se diferencia en cadenas de conidiosporos. Cada conidiosporo puede, a su vez, generar una colonia micelial. Otro gnero de inters es Nocardia cuyas colonias carecen de micelio areo o es escaso, con unos pocos conidiosporos en los extremos de las cortas ramas hifales o sin ellos, y finalmente las hifas se fragmentan totalmente en elementos bacilares (7). Thermoactinomyces, a diferencia de los otros gneros de actinomicetos, forma endosporos similares a los de Bacillus y Clostridium, en el extremo de pequeas ramificaciones (29). La figura 10 muestra un esquema de la morfologa de algunos actinomicetos. Nocardia, como la bacteria Mycobacterium, no se tie fcilmente por el colorante de Gram ni por otros, para colorearla se recurre al mtodo de Ziehl Neelsen donde el colorante (fucsina fenicada)
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
se aplica en caliente y con el fin de diferenciarla se decolora a los otros organismos con un cido mineral diluido. Esto se debe a que el pptidoglucano est unido a los arabino-galactanos esterificados con cidos miclicos de naturaleza cerosa (1).
1.2.4. Cianobacterias
Son organismos procariticos, unicelulares o filamentosos (por reunin de clulas individuales adheridas por sus extremos) que contienen, adems de la clorofila, un pigmento azulado llamado ficocianina. Estn presentes en suelos, y aguas dulces o saladas. A veces colonizan ambientes extremadamente inhspitos. Algunos convierten el nitrgeno atmsferico en compuestos que los vegetales aprovechan y fertilizan los campos y de manera muy especial los arrozales (1). Ciertas especies producen metabolitos secundarios txicos (por ej. un fosfato orgnico letal) que son liberados de las clulas en el tracto digestivo de los animales al beber agua con verdn (27). Algunas son unicelulares adheridas por un limo o dentro de una cpsula, ej. Gloeocapsa. Otras tambin unicelulares generan multiples beocitos en su interior, ej. Dermocarpa. Las que forman cadenas de clulas (tricomas) que pueden deslizarse, suelen tener clulas especiales donde ocurre la fijacin del nitrgeno molecular (heterocistos) y clulas de reposo (acinetos), ej. Nostoc, o carecer de ellas, ej. Oscillatoria. Unas pocas tienen una vaina alrededor del tricoma (12). La figura 11 presenta un esquema de la morfologa de cianobacterias. Los gneros Anabaena, Aphanizomenon, Microcystis y Nodularia contienen especies txicas (27). En cambio el gnero Spirulina es comestible y apreciado por el alto tenor de protenas, del orden del 50-60% del peso seco, con un aminograma similar al de la harina de soja. Las cianobacterias tambin pueden convivir en simbiosis con las plantas, debido a su capacidad de fijar el nitrgeno molecular, por ej. Anabaena en las cavidades dorsales de las hojas del helecho acutico Azolla, y Nostoc en los ndulos caulinares del arbusto tropical Gunnera (28).
1.2.5. Mixobacterias
Las mixobacterias son organismos sociales cuyas clulas son flexibles y no tienen una pared celular rgida, generalmente estn embebidas en un limo espeso. Al desplazarse segregan un material mucoso extracelular que se convierte en grandes avenidas por donde avanzan miles de clulas. El movimiento es muy coordinado. Cuando la poblacin emigra sobre el agar, se desplaza como una unidad indivisa. Incluso las especies que entran en letargo como esporos unicelulares, exhiben hbitos sociales durante buena parte de su ciclo vital. Muchas de ellas nunca se presentan aisladas, por el contrario, entran en una etapa de letargo en forma de cisto pluricelular que luego germina y libera una nueva poblacin de millares de mixosporos (29). Las mixobacterias son organismos del suelo y los cuerpos fructferos se encuentran sobre material vegetal en descomposicin o estircol. Algunas, como Myxococcus, forman cuerpos fructferos como gotitas con
10
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1 menos de 1 mm de dimetro. Otras, como Sporocytophaga forman adems de los bacilos delgados, unas clulas ovales como esporos llamadas microcistos. Pero Cytophaga no forma ni microcistos ni cuerpos fructferos (12). En el conjunto de la poblacin de mixobacterias se dejan sentir ondas pulstiles rtmicas. Las oleadas de mixobacterias que se acercan al centro o se alejan hacia el borde de la colonia en crecimiento, forman agregados en puntos especficos para construir los cistos o, en algunas gneros como Chondromyces, complejos cuerpos fructferos. La produccin de estas estructuras responde a cambios fsicos y nutricionales en el ambiente. Las clulas perciben estos cambios y transforman esta percepcin en una serie de hechos que implican agregacin, construccin del cuerpo fructfero pluricelular y la conversin de las clulas alargadas en mixosporos redondos, resistentes y metablicamente en reposo. Durante estos procesos monitorean la densidad celular, controlan el cronometrado de una serie de acontecimientos evolutivos, emprenden comportamientos tcticos, sufren autlisis evolutiva y se orientan en el espacio tridimensional (29). Las mixobacterias depredadoras se alimentan segregando enzimas en los huecos de las colonias para evitar su dilucin en el medio acuoso, que disuelven la cubierta celular externa de otros microorganismos. Cuando stos estallan, las mixobacterias absorben el contenido (22). La figura 12 muestra el diagrama del ciclo de vida de una mixobacteria (Myxococcus xanthus) .
1.3. Organismos eucariticos
Los cromosomas eucariticos estn asociados a una clase de protenas bsicas llamadas histonas, y experimentan movimientos en el momento de la divisin celular. Como en los procariotas, la informacin contenida en el ADN es transportada por los ARN mensajeros del ncleo hacia el citoplasma, donde esta informacin es descodificada y da lugar a la sntesis de las protenas en los ribosomas (14). Una clula eucaritica contiene orgnulos definidos por membranas que cumplen funciones esenciales, tales como las mitocondrias (produccin la energa), el retculo endoplsmico (sntesis de protenas en los ribosomas adheridos), el aparato de Golgi o dictiosoma (sntesis y almacenamiento de glicoprotenas), y en algas los cloroplastos (fotosntesis) (12). Las protenas son dirigidas hacia la ruta secretora por una secuencia terminal (seal pptido) que es reconocida por una partcula reconocedora de la seal. Despus de la unin del complejo ribosoma-partcula a la membrana del retculo endoplsmico, el traslado contina y el polipptido naciente llega al lumen del retculo endoplsmico donde una endoproteasa especfica quita la seal y, tiene lugar el ensamblaje (plegado y formacin de puentes disulfuro) y la N -glicosilacin de la protena. Despus las protenas dejan el retculo endoplsmico en vesculas y son transportadas al aparato de Golgi donde ocurren ulteriores modificaciones. De all salen las protenas para sus diversos destinos, otra vez en vesculas, hacia la membrana citoplasmtica (secrecin) o los lisosomas (digestin de molculas) (24). La mayor parte de la energa obtenida por la oxidacin de glucosa y cidos grasos es transformada en energa qumica utilizable por la clula mediante la sntesis del trifosfato de adenosina (ATP) en la cadena respiratoria. Estas reacciones ocurren en las mitocondrias. Tienen dos membranas, la interna est invaginada en crestas. Los pliegues contienen los componentes de la cadena de transporte de electrones y la ATP sintasa. Tambin un pequeo nmero de protenas es sintetizado en los ribosomas de la matriz mitocondrial, la que contiene tambin su propio ADN pues estos orgnulos estn dotados de la capacidad de multiplicarse (30). Todas las estructuras se desplazan por el citoplasma a lo largo de pequeos tubos huecos, los microtbulos, que son polmeros de tubulina nucleados por el centrosoma
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
11
(31). La organizacin de los microtbulos en flagelos y cilios, permite a algunos
eucariotas unicelulares nadar en el medio en que se encuentra (32).
1.3.1. Mohos
Los mohos se caracterizan por tener ncleo verdadero, carecer de pigmentos fotosintticos, poseer micelio con pared celular constituida por glucanos, quitosano y quitina (polmeros de glucosa, glucosamina y N-acetil-glucosamina, respectivamente) (7). En pocas ocasiones la pared est constituida enteramente de quitina. La pared celular del micelio de los hongos semeja un extenso sistema tubular por el que avanza protegido el citoplasma para su dispersin y bsqueda de nutrientes (1). Los elementos somticos tubulares que constituyen el micelio reciben el nombre de hifas. Las hifas pueden estar separadas en secciones, generalmente multinucleadas, por medio de septos perforados o bien carecer de ellos. Los hongos pueden reproducirse tanto sexual como asexualmente. Los hongos asexuales (anamorfos) generan varias clases de esporas asexuales por mitosis del ncleo celular (mitosporas) (33). La morfologa de las estructuras que contienen las esporas es muy variable y constituye una de las bases de la clasificacin de los hongos. El micelio somtico no es suficientemente discriminador para utilizarlo en la clasificacin. El color de muchos mohos que viven en la materia orgnica en descomposicin se debe al color de sus esporas asexuales. stas presentan varias tonalidades de color blanco, amarillo, azul, verde, rojo, pardo o negro (7). Los mohos generan varias clases de esporas asexuales, mono o pluricelulares. Las esporas se desarrollan en los esporforos, estructuras especializadas que se extienden en el aire a partir del micelio vegetativo, y las esporas se acumulan en el extreno superior de los mismos. Si las esporas estn encerradas en un esporangio (en forma de bolsa) se las llama esporangiosporas. Los conidios son esporas externas o sea no estn encerradas. Al madurar, estas esporas son esparcidas por el viento. Muchos mohos pueden reproducirse tambin a travs de esporas sexuales, generadas por meiosis, o divisin reductora, de un ncleo diploide (meiosporas). En la meiosis, el nmero de cromosomas se divide por la mitad. Las esporas sexuales contienen slo un cromosoma de cada par homlogo. La condicin diploide se restablece cuando dos estructuras haploides se unen, completando el ciclo vital. Los mohos a los que no se les conoce ciclo sexual, se consideran hongos imperfectos (33). Los mohos con estructuras reproductoras sexuales (teleomorfos) corresponden a tres grupos: ascomicetos, basidiomicetos y zigomicetos. Los ascomicetos producen sus esporas en ascos, que generalmente se forman dentro de un complejo cuerpo fructfero, el ascoma. De forma similar, los basidiomicetos desarrollan sus esporas sexuales externamente, en los basidios que se hallan en un complejo cuerpo fructfero: el basidioma. Pero este grupo tambin comprende a los carbones y las royas, organismos de inters agronmico por ser parsitos vegetales. Los zigomicetos producen zigosporas a veces visibles a ojo desnudo. En condiciones naturales, los mohos se reproducen en la mayora de los casos asexualmente, las estructuras reproductoras sexuales slo aparecen ocasionalmente en circunstancias favorables (33). La figura 13 muestra un moho (Rhizopus stolonifer).
12
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
1.3.2. Levaduras
La mayora de la numerosas especies de levaduras se han clasificado desde el punto de vista de la r eproduccin, que puede ser sexual o asexual. En la reproduccin vegetativa, generalmente, una clula madre da lugar a diversas clulas hijas por la formacin repetida de yemas en la superficie celular; en unas pocas levaduras la divisin asexual se hace por escisin celular luego de la duplicacin del ncleo (34). Tres grupos de hongos acogen a las levaduras: los ascomicetos, los basidiomicetos y los hongos imperfectos. El primer grupo incluye las levaduras cuyas estructuras reproductoras sexuales son os ascos sencillos que contienen ascosporas. Una clula l diploide de levadura sufre meiosis y forma de cuatro a ocho ascosporas, encerradas en el asco. Una ascospora es una clula haploide que al germinar genera una progenie de clulas haploides por mitosis (reproduccin asexual). Las clulas de diferente polaridad sexual se combinan para formar un nuevo organismo diploide. Entre las levaduras que pertenecen a los ascomicetos se encuentra Saccharomyces cerevisiae empleada para la fabricacin del pan y la fermentacin alcohlica. Los basidiomicetos agrupan un nmero reducido de levaduras. Los hongos imperfectos incluyen levaduras que se reproducen slo de forma asexual, ej. Candida tropicalis (7). En la figura 14 se muestra el aspecto de una levadura que se divide por escisin y forma meiosporas.
1.3.3. Macromicetos
Son hongos con cuerpos fructferos (ascoma o basidioma) que se miden en centmetros. Entre ellos son de inters los que forman asociaciones simbiticas con rboles (ectomicorrizas), por ej. Amanita, Boletus, Cortinarius, Lactarius, Russula, Scleroderma, Suillus, Tricholoma, y los que crecen sobre troncos descomponiendo la madera, por ej. Schyzophyllum, Polyporus, Coriolus, Xylaria (35). La figura 14 tambin muestra una seta venenosa.
1.3.4. Mixomicetos
Los mixomicetos son organismos eucariticos mucosos que forman plasmodio y luego un cuerpo fructfero de colores destacados, sobre troncos y hojas en descomposicin o postes viejos, de 0,5 a 1 cm (36). La asociacin celular en los hongos mucilaginosos est mediada por la interaccin entre protenas unidas a carbohidratos de una clula y receptores formados por oligosacridos especficos de otra. De este modo, la diferenciacin de los hongos mucilaginosos desde una forma somtica (unicelular) hasta la forma cohesiva (agregada) va acompaada de la aparicin de lectinas y glicoprotenas especficas en la superficie celular (17). Las esporas liberadas de los cuerpos fructferos germinan sobre una superficie hmeda y producen mixoflagelados que nadan o bien mixamebas. stos se alimentan de los nutrientes lquidos o por fagocitosis de bacterias, levaduras, esporas fngicas, etc. Las clulas flageladas pierden luego su flagelo y
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
13
entran en un estado ameboide. Estas clulas son mononucleadas. Ocasionalmente se fusionan en pares para dar mixozigotos. Estas amebas diploides se fusionan para dar un plasmodio (estructura multinucleada). Luego el plasmodio da origen al cuerpo fructfero o esporangio con numerosos esporas haploides (33). La figura 15 muestra las estructuras de un mixomiceto.
1.3.5. Protozoos
Los protozoos constituyen un grupo muy heterogneo de microorganismos eucariticos, unicelulares y mviles (con algunas excepciones). El ciclo de vida comprende dos fases: una de actividad, durante la cual se desplazan, nutren y reproducen; otra de reposo o enquistamiento. Se alimentan de bacterias, levaduras, algas y otros protozoos. Se los agrupan en: rizpodos que se desplazan por pseudpodos (amebas y testceos o foraminferos), flagelados con uno o ms flagelos, ciliados dotados de cilias y dos ncleos, y esporozoos parsitos. Se observa un esquema de los mismos en la figura 1 y 16. La poblacin de flagelados y amebas es del orden de 103 - 10 5 por gramo de suelo, la de ciliados 103/g y la de rizpodos testceos 10 3 - 104 /g. Participan en el equilibrio biolgico del suelo pues consumen grandes cantidades de bacterias, una ameba ingiere unas 40.000 bacterias entre cada divisin celular (4).
1.3.6. Algas
Son organismos fotosintticos eucariticos. L algas as del suelo viven en la proximidad inmediata a la superficie o sobre la misma. Predominan, arriba las diatomeas y abajo las clorofceas y xantofceas ubicuas. La humedad ptima es del 40 al 60% de la capacidad de retencin del agua por el suelo. Las algas libres o liquenizadas constituyen el estado inicial de la vegetacin rocas y suelos minerales infrtiles (4). En la figura 17 se muestra un alga roja del suelo.
1.3.7. Lquenes
Un lquen est constitudo por un hongo (el micobionte) junto con una o ms algas y/o cianobacterias (el fotobionte) viviendo en una relacin simbitica y formando un cuerpo estable e identificable (39). La figura 18 muestra una rama con varios lquenes.
1.4. Organismos acelulares
1.4.1. Virus
Un virus es un programa gentico que lleva de una clula a otra el mensaje "reprodceme". Consiste de un cido nucleico y una cubierta protenica conocida como
14
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1 cpside, a veces tambin tiene una envoltura. Hay muchas clases de virus y se los puede agrupar por el tipo de material gentico que portan. Algunos tienen una molcula de ARN monocatenario (cadena "ms") compuesto por varios miles de subunidades nucleotdicas, que puede ser ledo directamente por el aparato de traduccin del hospedador, el ribosoma, como si fuera un ARN mensajero propio. Un ejemplo de este tipo de virus es el del mosaico del tabaco. Otros virus, por ej. el de la rabia, cifran sus mensajes en cadenas "menos" de ARN, las que en el interior de la clula deben transcribirse en cadenas complementarias de tipo "ms" para que empiece la replicacin. Los retrovirus constituyen una tercera clase de virus de ARN monocatenario. Cuando un retrovirus infecta a una clula, la enzima retrotranscriptasa transforma la cadena de ARN vrico en ADN monocatenario y ste es copiado por la ADN-polimerasa celular a ADN bicatenario que puede integrarse al genoma del hospedador. Los reovirus, patgenos de plantas y animales, tienen ARN bicatenario. Ejemplos de virus con ADN monocatenario son los inovirus y con ADN bicatenario los miovirus, ambos patgenos de bacterias (39). Un fago (virus que infecta a bacterias) posee una sola molcula de cido nucleico que se inyecta en la clula bacteriana. La figura 19 muestra la forma de un tipo de bacterifago. En la figura 20 se observa el ciclo de un fago lambda con ADN bicatenario lineal cuyos extremos se unen una vez dentro de la bacteria y los genes dispuestos sobre ese anillo dirigen la sntesis de las protenas vricas, valindose de la maquinaria bacteriana de sntesis. Parte de las protenas son enzimas implicadas en la replicacin del cido nucleico del fago, tarea que realizan en conjuncin con enzimas de la propia bacteria. El virus se reproduce, es decir fabrica nuevas molculas de ADN y protenas de la cpsula. Los nuevos ADN se introducen en las cpsulas recin formadas y el hospedador acaba lisndose (40). Por otra parte, el fago puede insertarse en el ADN de la bacteria hospedadora. En el estado integrado, el ADN del fago es replicado junto con el ADN bacteriano y no se expresa la informacin contenida en el primero. Este fago no infeccioso, que puede pasar de clula en clula por herencia, se denomina profago. La excisin del fago en la clula lisognica le devuelve la virulencia. Otra forma de agrupar los virus es por la forma. Se llama nucleocpside al cido nucleico (o virin) con la cpside,
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
15
y puede estar desnuda o recubierta por una membrana. Una cpside consta de capsmeros, y en la mayora de los casos es simtrica. Se observan dos tipos de estructuras simtrcas: helicoidal e icosahdrica (12). En la figura 21 se muestra la forma de algunos virus.
1.4.2. Viroides
Los viroides son ms sencillos que los virus, pues son slo filamentos muy cortos de ARN. Producen enfermedades especficas de las plantas (42).
1.4.3. Priones
Se llama as a las protenas que actan como agentes infecciosos en algunas enfermedades del sistema nervioso de ovejas, vacas y otros animales (43), pero es discutible si se pueden considerar microorganismos.
1.7. Transferencia gentica en bacterias
En una bacteria la informacin gentica est codificada en una molcula de ADN larga y muy plegada para caber en la clula. El cromosoma bacteriano circular tiene al menos un milmetro de largo. La doble hlice formada por dos cadenas de nuclotidos consta de varios millones de pares de bases y la informacin total de la bacteria se despliega en un conjunto de varios miles de genes estructurales. En los procariotas y eucariotas la clave gentica y la bioqumica esencial de la transcripcin y la traduccin son las mismas, pero no as las seales de control. En las bacterias la informacin gentica codificada en un segmento contnuo de la doble hlice de ADN, que constituye el gen estructural, se transcribe directamente en un ARN mensajero cuya traduccin da lugar a una protena, generalmente una enzima (44). Antes de que una clula bacteriana se divida (transferencia vertical) ha de duplicarse su ADN (figura 22). Durante la replicacin del ADN se separan los pares de bases al alejarse ambas cadenas helicoidales. Las ADN polimerasas copian cada una de las hebras. Mientras se va construyendo la nueva hebra, sta se empareja con la que le sirve de molde. Pueden darse errores de replicacin por corrimiento cuando alguna de las hebras se desliza y establecen un emparejamiento inadecuado. En cada tanda de divisin bacteriana asexual habr alguna clula que porte una mutacin. La mutacin altera los genes de un microorganismo. La recombinacin reoordena los genes o parte de los mismos, y agrupa en un mismo individuo la informacin gentica de dos o ms organismos (44). Los tres mecanismos principales de intercambio horizontal de material gentico entre bacterias son: conjugacin, transformacin y transduccin. En todos estos procesos se transfiere ADN de la clula donadora a la receptora, pero difieren en la manera en que es transportado. La transferencia es seguida por la recombinacin y as el ADN de la
16
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1 bacteria donadora se integra al de la receptora. El intercambio de genes en las bacterias no est confinado dentro de una especie o especies afines, acontece entre bacterias de gneros o familias diferentes. La recombinacin homloga ocurre cuando los ADN que poseen secuencias de bases similares, se renen e intercambian segmentos mediante la rotura y ensamblaje de las cadenas. Otra forma de recombinacin que aade nuevas porciones al ADN que ya posee el microorganismo, es la especfica de sitio y requiere solo pequeos fragmentos de ADN homlogo para el reconocimiento. Cuando ambos ADN poseen una secuencia de reconocimiento se habla de recombinacin especfica de sitio doble, por ejemplo la insercin de un fago. Si solamente una molcula de ADN transporta esa secuencia la recombinacin especfica de sitio es simple y permite la insercin de una secuencia mediante la transposicin (12). Ambientes en los que ha observado transferencia gentica horizontal (41)
Ambientes Terrestres Acuticos
Transduccin Suelos, superficies de plantas Lagos, ocanos, ros, zonas de tratamiento de aguas residuales
Interior de organismos
Crustceos, ratones
Conjugacin Suelos, superficies de plantas Lagos, ocanos, sedimentos marinos, ros, epiliton (capas gelatinosas que recubren las piedras de los ros), zonas de tratamiento de aguas residuales Plantas, insectos, gallinas, ratones, humanos
Transformacin Suelos Sedimentos marinos, ros, epiliton de las piedras de los ros
Plantas, insectos, ratones
1.7.1. Conjugacin
Se descubri cuando se mezclaron mutantes nutricionales (auxotrficos) con diferentes requerimientos de la bacteria Gram-negativa Escherichia coli y crecieron sobre un medio mnimo siendo que, separadamente, necesitaban un suplemento de determinados nutrientes para poder multiplicarse. Los genes silvestres de una cepa completaron los genes mutados de la otra (figura 23). Este proceso requiere el contacto entre las clulas y se transfiere el factor sexual F (fertilidad) que es una molcula pequea de ADN circular de doble cadena (plsmido) desde la clula donadora (F+) a la receptora (F-). Las clulas donadoras tienen en la superficie los pelos de conjugacin (pili F) que sirven para facilitar el contacto (figura 24). La presencia de un plsmido altera la capacidad para sintetizar el pelo F y el desplazamiento del ADN para su transferencia a otra clula, adems modifica los receptores superficiales.
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
17
Slo unas pocas bacterias F+ pueden transferir ADN cromosomal. Son las que tienen el factor F integrado al cromosoma. Tales bacterias, llamadas Hfr, transfieren sus genes con una frecuencia muy alta. Durante la transferencia el ADN bacteriano se replica a partir del punto de insercin del factor F y la cadena recin formada entra en la clula aceptora. Este proceso es seguido de la recombinacin homloga del ADN de ambas clulas dentro de la bacteria receptora (figura 25). La introduccin de unas cuantas bacterias con plsmidos en una poblacin receptora, puede convertirla en portadora de plsmidos en poco tiempo (45). En las bacterias Gram-positivas la conjugacin no est mediada por pili. Antes del intercambio gentico, la clula receptora secreta substancias que estimulan la sntesis de protenas aglutinantes por las potenciales donadoras. Una vez que las bacterias se asocian, se forman los poros necesarios para la transferencia del ADN (41).
1.7.2. Transformacin
Algunas bacterias pueden captar ADN desnudo, por ejemplo las clulas de una colonia rugosa de Streptococcus pneumoniae al ser mezcladas con el ADN de una cepa virulenta con colonias lisas adquieren la cualidad de producir colonias lisas. La transformacin suele ocurrir tanto en bacterias Gram-positivas como en Gram-negativas, y es utilizada para introducir genes extraos en una bacteria (12). Un fragmento de ADN se puede insertar en un plsmido pequeo especializado conocido como vector de clonacin usando una enzima especfica. Este ADN recombinante se usa para transformar las bacterias que crecern luego en un medio selectivo apropiado para detectarlas. La capacidad de aislar ADN de un organismo e inducir su incorporacin en otro no relacionado constituye el fundamento de una de las manipulaciones del ADN recombinante (figura 26). El plsmido sirve as de vector para genes que no tienen equivalente en el organismo receptor y que, por lo tanto, no podran heredarse de forma estable mediante recombinacin homloga. Estos genes pueden as transmitirse a travs de generaciones sucesivas conforme el plsmido va replicndose (44). Pero a veces la bacteria pierde el plsmido espontneamente. La mayora de las especies de rizobios, bacterias de gran importancia agronmica, contienen plsmidos con la informacin gentica para la simbiosis en leguminosas (46).
1.7.3. Transduccin
Los genes bacterianos se pueden transferir de una cepa a otra usando bacterifagos como vectores (figura 19), pero este fenmeno no ocurre en todas las especies de bacterias. Segn el comportamiento del bacterifago la transduccin puede ser:
18
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
a) general o inespecfica
Comnmente el bacterifago se replica dentro de la clula que infecta y con la ruptura de la misma se liberan las nuevas partculas vricas. Estas partculas infectan otras clulas completando el ciclo de lisis. Ocasionalmente durante el proceso ltico el genoma bacteriano se fragmenta y algunos segmentos pasan a formar parte del nuevo bacterifago. stos suelen carecer de parte de su propio genoma, pero aunque defectuosos son capaces de infectar nuevas bacterias y dejarles la porcin de ADN bacteriano que transportan. Este ADN suele recombinarse con el genoma de la clula infectada (figura 27). La porcin de genoma bacteriano transportada por bacterifagos es por lo general menor que 1% del ADN total. La mayora de las partculas vricas son normales y no participan en la transduccin (12). Los bacterifagos moderados tambin son capaces de formar una relacin estable con sus hospedadores. Luego de la infeccin suelen incorporarse al genoma bacteriano como profago. ste suele codificar otras funciones adems de las especficas del bacterifago, por ejemplo la produccin de toxinas. La bacterias que albergan profagos se llaman lisognicas (figura 20). Los profagos suelen vivir en armona con su hospedador hasta que una situacin de stress ambiental induce el ciclo ltico. Cuando el bacterifago se escinde del genoma bacteriano puede acarrear una porcin de este ltimo. Estos fagos defectuosos invaden nuevas bacterias. La integracin al genoma de la clula hospedadora es esencial para la trasferencia de los genes. Como cada profago tiene un particular lugar de insercin en el genoma de la bacteria, solamente los
b) especializada
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
19
genes adyacentes pueden ser transferidos por este proceso (12 ). La transposicin es la recombinacin que presentan los transposones o las secuencias de insercin. Las secuencias de ADN cambian de lugar en el genoma y se insertan en una regin que no guarda homologa con ella. Si esta integracin sucede dentro de un gen se produce una mutacin, pues se rompe la continuidad de la informacin codificada (47).
1.7.4. Fusin de protoplastos
Las barreras naturales a la recombinacin entre organismos diferentes pueden romperse a menudo mediante la preparacin de protoplastos, clulas bacterianas cuyas paredes externas se han eliminado poniendo al descubierto la membrana celular. Como las membranas celulares poseen aproximadamente la misma composicin en la mayora de los organismos, puede inducirse la fusin de protoplastos de especies distintas, formando un clula hbrida en la que los genes quedan expuestos a la recombinacin (figura 28). Tambin es una tcnica eficaz para aumentar la frecuencia de recombinacin intraespecfica en los que el apareamiento natural es raro. La operacin se lleva a cabo en una solucin cuya presin osmtica equilibra la presin interior de las clulas, para que no estalle la membrana celular. Despus se induce la regeneracin de la pared de los protoplastos hbridos y se obtiene un cultivo normal (44).
Referencias
1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. Madigan TM, Martinko JM, Parker J. Brock-Biology of Microorganisms. Prentice Hall, Upper Saddle River, NJ, 2003 Stanier R.Y. et al. Microbiologa. 4 ed. Barcelona, Revert. 1984 Fenchel T, King GM, Blackburn TH. Bacterial Biogeochemistry. 2 ed. Academic Press, San Diego, 2000, cap.1 Dommergues Y. & Mangenot F. cologie microbienne du sol. Paris, Masson et Cie. 1970, pp. 18, 27, 36, 48-69 Raibaud P. & Ducluzeau R. Las bacterias del tubo digestivo. Mundo Cientfico 34: 292-302, 1984 Potera C. Biofilms invade microbiology. Science 273: 1795 -1797, 1996 Phaff H.J. Microorganismos industriales. Investigacin y Ciencia 62: 22-37, 1981 Woese C.R. Archibacterias. Investigacin y Ciencia 59: 48 -61, 1981 Margulis L.& Sagan D. El origen de las clulas eucariontes. Mundo Cientfico 46: 366 -374, 1985 Doolittle WF. Nuevo rbol de la vida. Investigacin y Ciencia 283: 26-32, 2000 Hawksworth D.L. et al. Dictionary of the Fungi. Wallingford, CAB International 1995, pp. 48, 228, 442 Schlegel H.G. General microbiology. Cambridge University Press 1993, pp. 89 -169 Duve C. El origen de las clulas eucariotas. Investigacin y Ciencia 237: 18-26, 1996 Flix M.A. & Karsenti E. La divisin celular. Mundo Cientfico 154: 132-140, 1995 Mahon C.R. & Manuselis G. Texbook of Diagnostic Microbiology. Philadelphia, W.B.Saunders Co. 1995, p.9 Rietschel ET, Brade H. Endotoxinas bacterianas. Investigacin y Ciencia 193: 16 24, 1992 Bretscher M.S. Molculas de la membrana celular. Investigacin y Ciencia 111: 66-75, 1985 Ofek I & Sharon N. Cmo se adhieren las bacterias a las clulas. Mundo Cientfico 25: 564 - 566, 1983 Sra M. Conserved anchoring mechanisms between crystalline cell surface S -later proteins and secondary cell wall polymers in Gram positive bacteria? Trends in Microbiology 9: 47-49, 2001 Novick R.P. Plsmidos. Investigacin y Ciencia 53: 46-59, 1981 Nieto-Jacobo MF, Guevara-Garca A, Herrera-Estrella L. Plantas transgnicas. Investigacin y Ciencia 268: 70-80, 1999 Shapiro J.A. Las bacterias, organismos pluricelulares. Investigacin y Ciencia 143: 56-64, 1988
20
Leonor Carrillo. 2003. Microbiologa Agrcola. captulo 1
23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. 35. 36. 37. 38. 39. 40. 41. 42. 43. 44. 45. 46. 47. Mller-Jensen J, Jensen RB, Gerdes K. Plasmid and chromosome segregation in prokaryotes. Trends in Microbiology 8:313-320, 2000 Becker B. & Melkonian M. The secretory pathway of protists. Microbiological Reviews 60: 697-721, 1996 Bernander R. Chromosome replication, nucleoid segregation and cell divisin in Archaea. Trends in Microbiology 8: 278-283, 2000 Williams S.T. et al. Bergeys Manual of Systematic Bacteriology. Volume 4.Baltimore, Williams & Wilkins 1989, p.2341 Carmichael W.W. Toxinas de cianobacterias. Investigacin y Ciencia 210: 22-29, 1994 Dommergues Y et al. Fijacin del nitrgeno t agricultura tropical. Mundo Cientfico 45: 276 285, 1985 Dworkin M. Social and developmental biology of the myxobacteria. Microbiological Reviews 60: 70-102, 1996 Durand R. El acoplamiento de las mitocondrias. Mundo Cientfico 57: 376-385, 1986 Glover D.M. et al. El centrosoma. Investigacin y Ciencia 203: 22 -29, 1993 Porter K.R. & Tucker J.B. El armazn celular. Investigacin y Ciencia 56: 16-28, 1981 Webster J. Introduction to fungi. 2ed. Cambrige, Cambridge University Press, 1986 Gaillardin C. & Heslot H. La levadura. Mundo Cientfico 71: 716-727, 1987 Hudson H.J. Fungal biology. London, Edward Arnold 1986, pp.99-102, 184 Ageitos Z.J. & Lopretto E.C. Los invertebrados. Tomo I. Buenos Aires, EUDEBA. 1983, pp. 104, 136, 163, 294 Lee R.E. Phycology. New York, Cambridge University Press 1980,pp.175,205,429 Purvis W, Wedin M. El xito todo terreno de los lquenes. Mundo Cientfico 200: 56 59, 1999 Eigen M. Cuasiespecies vricas. Investigacin y Ciencia 204: 14-22, 1993 Stahl F.W. Recombinacin gentica. Investigacin y Ciencia 127: 42-55, 1987 Miller RV. Intercambio de genes bacterianos en la naturaleza. Investigacin y Ciencia 258: 1318, 1998 Diener TO. Viroides. Investigacin y Ciencia 54: 18 26, 1981 Eigen M. Priones y encefalopata espongiforme bovina. Investigacin y Ciencia 298: 7483, 2001 Hopwood DA. Programacin gentica de microorganismos industriales. Investigacin y Ciencia 62, 40-54, 1981 Moxon ER, Wills C. Microsatlites de ADN. Investigacin y Ciencia 270: 68 74, 1999 Martnez E, Palacios R, Mors J. Cepas mejoradas de Rhizobium. Investigacin y Ciencia 265: 1419, 1998 Sobern G. Mecanismo de nodulacin de las leguminosas. Investigacin y Ciencia 103: 613, 1985
You might also like
- Cultivo de Banano GuatemalaDocument27 pagesCultivo de Banano GuatemalaXavier L Mipi100% (1)
- Análisis funcional y microbiológico de derivados lácteos y cárnicosFrom EverandAnálisis funcional y microbiológico de derivados lácteos y cárnicosNo ratings yet
- Microbiologia Industrial LibroDocument108 pagesMicrobiologia Industrial LibroAlejandro Canaza Jorges0% (1)
- Los inicios de la vida: La evolución en la Tierra precámbricaFrom EverandLos inicios de la vida: La evolución en la Tierra precámbricaNo ratings yet
- Hongos y LevadurasDocument24 pagesHongos y LevadurasL C100% (13)
- Resumen Y Guía De Estudio - 10% Humano: Los Microbios Que Habitan En Su Cuerpo Son La Clave Para La Salud Y La FelicidadFrom EverandResumen Y Guía De Estudio - 10% Humano: Los Microbios Que Habitan En Su Cuerpo Son La Clave Para La Salud Y La FelicidadRating: 1 out of 5 stars1/5 (1)
- Características Generales de Las BacteriasDocument16 pagesCaracterísticas Generales de Las BacteriasEstrella Jatziry Espíndola GuerreroNo ratings yet
- Analisis de Orina InterpretacionDocument5 pagesAnalisis de Orina InterpretacionQUIMICO CLINICO WILLIANS SANCHEZ95% (20)
- Resumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSFrom EverandResumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSNo ratings yet
- Bacterias Gram NegativasDocument27 pagesBacterias Gram Negativascesar alexisNo ratings yet
- Guias para La Calidad Del Agua Potable OMSDocument408 pagesGuias para La Calidad Del Agua Potable OMSRomulo Aycachi Inga80% (5)
- Anatomía y Fisiología de Las Células ProcariotasDocument20 pagesAnatomía y Fisiología de Las Células ProcariotasLis Pérez25% (4)
- Hoja de PresentaciónDocument11 pagesHoja de PresentaciónKmlo SaChezNo ratings yet
- Evolución de La Celula Procariota y EucariotaDocument7 pagesEvolución de La Celula Procariota y EucariotaAngely Rojas ContrerasNo ratings yet
- Ciano BacteriasDocument16 pagesCiano BacteriasPoolAndersonPachecoCastilloNo ratings yet
- Célula ProcariotaDocument4 pagesCélula ProcariotaAnonymous hrOP69oNdcNo ratings yet
- Microbiologia ApuntesDocument54 pagesMicrobiologia ApuntesAgustin HumerezNo ratings yet
- Taller Teoria EndosimbioticaDocument7 pagesTaller Teoria EndosimbioticaCarlos De la OssaNo ratings yet
- Célula Procariota - Wikipedia, La Enciclopedia LibreDocument14 pagesCélula Procariota - Wikipedia, La Enciclopedia LibreSolaine TovarNo ratings yet
- Bio Lab Prac4Document13 pagesBio Lab Prac4MaJo MartinezNo ratings yet
- Estudio DirigidoDocument18 pagesEstudio DirigidoLauraNo ratings yet
- Reinos Celula DigstivoDocument11 pagesReinos Celula DigstivoAndres r1No ratings yet
- CLASE UNIDAD II - PPSXDocument111 pagesCLASE UNIDAD II - PPSXfernandohualNo ratings yet
- Cuantos ReinosDocument8 pagesCuantos ReinosAndrea LiconaNo ratings yet
- Bacterias tipos, morfología y su importanciaDocument21 pagesBacterias tipos, morfología y su importanciaLeydi Lisbet ParedesNo ratings yet
- El Reino Monera o Mónera Esta Formado Por Las BacteriasDocument12 pagesEl Reino Monera o Mónera Esta Formado Por Las BacteriasManuel MontillaNo ratings yet
- Práctica 7 La CélulaDocument5 pagesPráctica 7 La CélulaJuan HernándezNo ratings yet
- Celula EucariotaDocument5 pagesCelula EucariotaluciaNo ratings yet
- ProcariotaDocument42 pagesProcariotaNotSrPaltaNo ratings yet
- La Célula Procariota Los Procariotas Son El Grupo Más Antiguo de Organismos Sobre La Tierra y Los Más Abundantes en EllaDocument3 pagesLa Célula Procariota Los Procariotas Son El Grupo Más Antiguo de Organismos Sobre La Tierra y Los Más Abundantes en EllaValeria Hernandez ChavezNo ratings yet
- Grupos Microbianos PDFDocument36 pagesGrupos Microbianos PDFsilviaNo ratings yet
- Reino MoneraDocument7 pagesReino MoneraHarol Anthony Mines ManosalvaNo ratings yet
- Citologia Parte IiDocument5 pagesCitologia Parte IiLiiseth NarvaezNo ratings yet
- Organismos unicelulares y pluricelularesDocument20 pagesOrganismos unicelulares y pluricelularesdiegoalexanderpancheNo ratings yet
- Organelos CitoplasmáticoDocument16 pagesOrganelos CitoplasmáticoAlexandra Bernedo UndaNo ratings yet
- Celula ProcariotaDocument8 pagesCelula ProcariotaJUAN ALEXANDER YOC MENDEZNo ratings yet
- CELULADocument16 pagesCELULALAURA VIRTUDESNo ratings yet
- Unidad III BACTERIAS Y ARQUEAS DBDocument12 pagesUnidad III BACTERIAS Y ARQUEAS DBRita RodríguezNo ratings yet
- Reino MoneraDocument16 pagesReino Moneragermain figueroaNo ratings yet
- Bacteria DefinicionDocument12 pagesBacteria DefinicionYunuen Mendoza CalderonNo ratings yet
- Universidsad Autonoma Gabriel Rene MorenoDocument5 pagesUniversidsad Autonoma Gabriel Rene Morenoromeroisis41No ratings yet
- Microbiologia BacteriasDocument16 pagesMicrobiologia BacteriasSol VillagraNo ratings yet
- Reino MoneraDocument6 pagesReino MoneraMiguel De la cruzNo ratings yet
- Qué Es El Reino MoneraDocument9 pagesQué Es El Reino Moneradelacruzs19No ratings yet
- Clase 24-01 La Ciencia de La MicrobiologíaDocument6 pagesClase 24-01 La Ciencia de La MicrobiologíaMace AsheNo ratings yet
- El Origen de Las Células EucariontesDocument16 pagesEl Origen de Las Células EucariontesMacarena FranchiNo ratings yet
- Casio FX 3600p ManualDocument13 pagesCasio FX 3600p ManualAlfredoNo ratings yet
- Resumen ParcialDocument12 pagesResumen ParcialZ I.No ratings yet
- 4.2.1 ArqueobacteriasDocument4 pages4.2.1 ArqueobacteriasAlex OrtizNo ratings yet
- 18 Clasificacion MicroorganismosDocument7 pages18 Clasificacion Microorganismosttonitta6956No ratings yet
- CITOLOGIADocument25 pagesCITOLOGIAJosue CruzNo ratings yet
- REINO MONERA - GuíaDocument17 pagesREINO MONERA - GuíaNadia N. Salattino50% (2)
- Actividad 1 Celulas Eucariotas.Document19 pagesActividad 1 Celulas Eucariotas.Charly Romero SuarezNo ratings yet
- Procariotas y EucariotasDocument27 pagesProcariotas y EucariotasChristian YungaNo ratings yet
- edeDocument7 pagesedeelimarNo ratings yet
- Lab N°2 Marco Teórico Observación de Bacterias, Hongos, Protozoos y Célula Animal BIOLOGIADocument6 pagesLab N°2 Marco Teórico Observación de Bacterias, Hongos, Protozoos y Célula Animal BIOLOGIAPato Carmona CortésNo ratings yet
- Los 5 ReinosDocument7 pagesLos 5 Reinos23418872No ratings yet
- Lab N°2 Marco Teórico Observación de Bacterias, Protozoos y Célula Animal KINEDocument6 pagesLab N°2 Marco Teórico Observación de Bacterias, Protozoos y Célula Animal KINEcamila_munozleal100% (2)
- Modulo de Ciencias NaturalesDocument20 pagesModulo de Ciencias Naturalesyoamna bermudezNo ratings yet
- Clase 1Document19 pagesClase 1Remigio BàezNo ratings yet
- Seminario I. Células ProcariotasDocument22 pagesSeminario I. Células Procariotasmaria teresa zuluagaNo ratings yet
- Edición Especial: Año II - #1232Document111 pagesEdición Especial: Año II - #1232Mayuri EscobarNo ratings yet
- Genotipós SigatokaDocument13 pagesGenotipós SigatokaXavier L MipiNo ratings yet
- Cuadro Nacional de Medicamentos QUE SI CUBREDocument896 pagesCuadro Nacional de Medicamentos QUE SI CUBREKatherine BarzalloNo ratings yet
- Solución de Caso - DRH Brasil - Cosmopolita - 5%Document3 pagesSolución de Caso - DRH Brasil - Cosmopolita - 5%Xavier L Mipi100% (1)
- Sogatoka en La Hoja PDFDocument8 pagesSogatoka en La Hoja PDFXavier L MipiNo ratings yet
- Prosp Pharma Analytik ES WebDocument28 pagesProsp Pharma Analytik ES WebFrancisco ChavezNo ratings yet
- La HospitalidadDocument33 pagesLa HospitalidadNadyeli Gutiérrez Solis100% (1)
- La HospitalidadDocument33 pagesLa HospitalidadNadyeli Gutiérrez Solis100% (1)
- Contenido Modulo II Servicio Al Cliente PDFDocument40 pagesContenido Modulo II Servicio Al Cliente PDFcamilo_lozada-180% (5)
- Bases de Microbiología IndustrialDocument1 pageBases de Microbiología IndustrialXavier L MipiNo ratings yet
- Guia de Equivalentes TerapeuticosDocument12 pagesGuia de Equivalentes TerapeuticosXavier L MipiNo ratings yet
- Química orgánica: Aromaticidad PMODocument110 pagesQuímica orgánica: Aromaticidad PMOCalamidad Loco PeligroNo ratings yet
- Contenido Modulo II Servicio Al Cliente PDFDocument40 pagesContenido Modulo II Servicio Al Cliente PDFcamilo_lozada-180% (5)
- Determinacion de Agua 2014Document21 pagesDeterminacion de Agua 2014Xavier L MipiNo ratings yet
- Tecnologia FarmaceuticaDocument9 pagesTecnologia FarmaceuticaXavier L MipiNo ratings yet
- Malleus Maleficarum (Segunda Parte)Document82 pagesMalleus Maleficarum (Segunda Parte)Enigmatico100% (1)
- Alergia Elemental - Javier FernandezDocument185 pagesAlergia Elemental - Javier FernandezEliazar SuarezNo ratings yet
- Monografia Plátano2010Document23 pagesMonografia Plátano2010Alfonso Torres De la ONo ratings yet
- APUNTE Micologia GeneralDocument6 pagesAPUNTE Micologia Generalronal1800No ratings yet
- Guia-Stata7 - Datos de Panel PDFDocument10 pagesGuia-Stata7 - Datos de Panel PDFjanmferNo ratings yet
- Fichas Imagenes de EnfermedadesDocument64 pagesFichas Imagenes de EnfermedadesXavier L MipiNo ratings yet
- Geoquimica Recreativa - Alexandr FersmanDocument541 pagesGeoquimica Recreativa - Alexandr FersmanMarisela Amaya VanegasNo ratings yet
- Ley de BeerDocument10 pagesLey de Beermedun2009No ratings yet
- Manual de Parasitologia Ins PDFDocument101 pagesManual de Parasitologia Ins PDFAnonymous Ru0zFT6T100% (1)
- HepatitisDocument41 pagesHepatitisXavier L MipiNo ratings yet