You might also like
- Autismo e Neuronio Espelho PDFDocument6 pagesAutismo e Neuronio Espelho PDFElizete E ErildoNo ratings yet
- Sensação Percepção PropriocepçãoDocument3 pagesSensação Percepção PropriocepçãoDouglas TeixeiraNo ratings yet
- Pensamento e LinguagemDocument176 pagesPensamento e LinguagemJussara BarbosaNo ratings yet
- Inteligência Sensório-MotorDocument11 pagesInteligência Sensório-Motorvera_viegas100% (2)
- Atenção seletiva e ilusões perceptivasDocument26 pagesAtenção seletiva e ilusões perceptivasmorghana de oliveiraNo ratings yet
- Aula 01 - Fonética ArticulatóriaDocument19 pagesAula 01 - Fonética ArticulatóriaGeysa Andrade Da SilvaNo ratings yet
- As Funções Mentais Superiores - DELARI JRDocument6 pagesAs Funções Mentais Superiores - DELARI JRMarcelo Maia CirinoNo ratings yet
- Afasia de Condução: Substratos AnatômicosDocument55 pagesAfasia de Condução: Substratos AnatômicospatriciasusckingNo ratings yet
- A aula expositiva dialogada em uma perspectiva freireanaDocument13 pagesA aula expositiva dialogada em uma perspectiva freireanaEduardo Dias PereiraNo ratings yet
- A Formulação Do Conceito de Homeostase Por Walter CannonDocument15 pagesA Formulação Do Conceito de Homeostase Por Walter Cannonroberto-martinsNo ratings yet
- Neurocincia Cognitivae Desenvolvimento HumanoDocument23 pagesNeurocincia Cognitivae Desenvolvimento HumanoRAFAEL RAMOSNo ratings yet
- Desenvolvimento cognitivo segundo PiagetDocument6 pagesDesenvolvimento cognitivo segundo PiagetIsolde KozowskiNo ratings yet
- FREUD - Desenvolvimento Da Infância e AdolescênciaDocument26 pagesFREUD - Desenvolvimento Da Infância e AdolescênciaLeti MouraNo ratings yet
- Funções Mentais SuperioresDocument21 pagesFunções Mentais SuperioresPedro Paulo Valadão100% (2)
- O QUE É SER SURDO E OUVINTEDocument12 pagesO QUE É SER SURDO E OUVINTEkmillabNo ratings yet
- Sintaxe da Libras e Gramática GerativaDocument24 pagesSintaxe da Libras e Gramática GerativaO que diz a Bíblia?No ratings yet
- As Funções Mentais Superiores 1Document29 pagesAs Funções Mentais Superiores 1Kelyo Almeida100% (1)
- Aula 9 - Texto Ato ReflexoDocument3 pagesAula 9 - Texto Ato ReflexoKeyla SilvaNo ratings yet
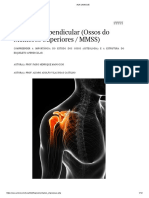
- Esqueleto ApendicularDocument12 pagesEsqueleto ApendicularMarcelo MarcosNo ratings yet
- Vigotski e o Aprender A Aprender Critica As AproprDocument3 pagesVigotski e o Aprender A Aprender Critica As AproprBruna OliveiraNo ratings yet
- Biomecânica PosturalDocument12 pagesBiomecânica PosturalFlávio MartinsNo ratings yet
- O Sistema LímbicoDocument14 pagesO Sistema LímbicoAngelita ValcanaiaNo ratings yet
- Fonética Articulatória: Descrição dos Sons do Português BrasileiroDocument19 pagesFonética Articulatória: Descrição dos Sons do Português BrasileiroJuan Carlos MPNo ratings yet
- LEONTIEV Alexei N. - Sobre o Desenvolvimento Criativo de VigotskiDocument28 pagesLEONTIEV Alexei N. - Sobre o Desenvolvimento Criativo de VigotskiLeonardo MorallesNo ratings yet
- Slides Bases Neurobiológicas Da LinguagemDocument29 pagesSlides Bases Neurobiológicas Da LinguagemAdriana Dettmann100% (1)
- Emoções - William JamesDocument1 pageEmoções - William JamesJoana Inês PontesNo ratings yet
- Psicanálise e neuroplasticidade: estimulando a neurogênese hipocampalDocument2 pagesPsicanálise e neuroplasticidade: estimulando a neurogênese hipocampaljrtdl573No ratings yet
- Como o sono afeta a aprendizagemDocument14 pagesComo o sono afeta a aprendizagemFábio RossetoNo ratings yet
- Apostila Transtornos de Aprendizagem X DesenvolvimDocument23 pagesApostila Transtornos de Aprendizagem X DesenvolvimPatriciaNo ratings yet
- Introdução à Psicomotricidade emDocument38 pagesIntrodução à Psicomotricidade emStefanny Schmitz100% (1)
- Quadro Sintese de Piaget Vygotsky WallonDocument5 pagesQuadro Sintese de Piaget Vygotsky WallonMauricio Vieira S. Pinto100% (1)
- Exercícios neurofuncionaisDocument14 pagesExercícios neurofuncionaisAndréa Frazão100% (1)
- Resumo de NeuropsicologiaDocument4 pagesResumo de NeuropsicologiaCamillaFreireNo ratings yet
- Desenvolvimento da LinguagemDocument70 pagesDesenvolvimento da LinguagemVanessa VolpattoNo ratings yet
- Estudo Da Supressão Do Comportamento Verbal Da GagueiraDocument18 pagesEstudo Da Supressão Do Comportamento Verbal Da GagueiraBianca GarciaNo ratings yet
- A Construção Da SubjetividadeDocument8 pagesA Construção Da SubjetividadeMarcus NunesNo ratings yet
- Método Beuttenmüller além das palavrasDocument10 pagesMétodo Beuttenmüller além das palavrasCamila Rabelo100% (1)
- Ensinando leitura e produção textual nos anos iniciaisDocument47 pagesEnsinando leitura e produção textual nos anos iniciaisAurismar Ferreira De Sousa SousaNo ratings yet
- A Evolução Do Grafismo e A Escrita Alfabética em Crianças de Dois A Seis AnosDocument20 pagesA Evolução Do Grafismo e A Escrita Alfabética em Crianças de Dois A Seis AnosUnicidade do Conhecimento100% (9)
- O Papel das Emoções na Tomada de Decisão segundo António DamásioDocument11 pagesO Papel das Emoções na Tomada de Decisão segundo António DamásioEliane FernandesNo ratings yet
- Relação indivíduo-comunidade em A OndaDocument6 pagesRelação indivíduo-comunidade em A OndaJuh FerrariNo ratings yet
- Teorias da Aquisição da LinguagemDocument55 pagesTeorias da Aquisição da LinguagemMariana Correia100% (1)
- Filosofia e Ética UbuntuDocument4 pagesFilosofia e Ética UbuntuFatima Nazareth100% (1)
- Música e estados alterados de consciênciaDocument11 pagesMúsica e estados alterados de consciênciaadrianaalmeidasNo ratings yet
- Psicomotricidade na educaçãoDocument14 pagesPsicomotricidade na educaçãoFelipe OliveiraNo ratings yet
- Observação da interação social de um portador de TEA na série AtypicalDocument7 pagesObservação da interação social de um portador de TEA na série AtypicalCris LopesNo ratings yet
- Os principais tipos de sons da língua portuguesaDocument7 pagesOs principais tipos de sons da língua portuguesaFelipe de SouzaNo ratings yet
- A Musicoterapia Nas Dificuldades de AprendizagemDocument3 pagesA Musicoterapia Nas Dificuldades de AprendizagemDébora Dos Santos100% (1)
- Engrama Cerebral - PesquisaDocument4 pagesEngrama Cerebral - PesquisaLeonardo Do CarmoNo ratings yet
- Apostila Educ PsicomotoraDocument52 pagesApostila Educ PsicomotoraAmfab100% (2)
- Pensamento e linguagem convergem e divergemDocument29 pagesPensamento e linguagem convergem e divergemJana Henning50% (2)
- Aula 3 Texto 2 PiagetDocument5 pagesAula 3 Texto 2 Piagetsophiosca5745No ratings yet
- Percorrer a Cidade a Pé: Ações Teatrais e Performativas no Contexto UrbanoFrom EverandPercorrer a Cidade a Pé: Ações Teatrais e Performativas no Contexto UrbanoNo ratings yet
- Neurônio EspelhoDocument11 pagesNeurônio EspelhoRenata Moraes RibeiroNo ratings yet
- Neurônios EspelhoDocument2 pagesNeurônios EspelhoJhonnata Pereira100% (1)
- Neurônios EspelhoDocument6 pagesNeurônios EspelhoMarcos MoscatelliNo ratings yet
- Neuronios EspelhoDocument5 pagesNeuronios EspelhoKarin SantosNo ratings yet
- Neurônios-espelho, empatia e neuromoralidadeDocument17 pagesNeurônios-espelho, empatia e neuromoralidadeCidinha LimaNo ratings yet
- Bases biológicas da atençãoDocument5 pagesBases biológicas da atençãobolctbaNo ratings yet
- Ginastica Laboral PDFDocument9 pagesGinastica Laboral PDFroggerio38souzaNo ratings yet
- A Influencia Do Estagio Extracurricular Cadernos Educacao PDFDocument9 pagesA Influencia Do Estagio Extracurricular Cadernos Educacao PDFMarcelo Peixoto SouzaNo ratings yet
- Código de Ética dos Profissionais de Educação FísicaDocument8 pagesCódigo de Ética dos Profissionais de Educação FísicaMarcelo Peixoto SouzaNo ratings yet
- Exame clínico odontológico: sinais, sintomas e anamneseDocument9 pagesExame clínico odontológico: sinais, sintomas e anamneseDiana DiasNo ratings yet
- Docencia OnlineDocument12 pagesDocencia OnlineMarcelo Peixoto SouzaNo ratings yet
- Saúde do Trabalhador: Ginástica Laboral e seus BenefíciosDocument37 pagesSaúde do Trabalhador: Ginástica Laboral e seus BenefíciosMarcelo Peixoto SouzaNo ratings yet
- Cinesioterapia: exercícios terapêuticos na fisioterapiaDocument3 pagesCinesioterapia: exercícios terapêuticos na fisioterapiaffelipee100% (1)
- Diretrizes da Educação Inclusiva no Estado da BahiaDocument148 pagesDiretrizes da Educação Inclusiva no Estado da BahiaCamilla BuergerNo ratings yet
- CIF e A FisioterapiaDocument7 pagesCIF e A FisioterapiaMarcelo Peixoto SouzaNo ratings yet
- Importância da Sociologia na formação de professores de Educação FísicaDocument6 pagesImportância da Sociologia na formação de professores de Educação FísicaMarcelo Peixoto SouzaNo ratings yet
- Revista FAEEBA N41 PDFDocument260 pagesRevista FAEEBA N41 PDFMarcelo Peixoto Souza100% (1)
- A Universidade e A Sociedade Do ConhecimentoDocument14 pagesA Universidade e A Sociedade Do ConhecimentoMarcelo Peixoto SouzaNo ratings yet
- Conceito de AcessibilidadeDocument2 pagesConceito de AcessibilidadeMarcelo Peixoto SouzaNo ratings yet
- Fisioterapia Estática e Classificação dos MúsculosDocument19 pagesFisioterapia Estática e Classificação dos MúsculosMarcelo Peixoto SouzaNo ratings yet
- Metodo Mezieres PDFDocument19 pagesMetodo Mezieres PDFGigiEnergieNo ratings yet
- Cinesioterapia: exercícios terapêuticos na fisioterapiaDocument3 pagesCinesioterapia: exercícios terapêuticos na fisioterapiaffelipee100% (1)
- 395 Trabalho Muscular 1mDocument11 pages395 Trabalho Muscular 1mRafael MathiasNo ratings yet
- Artigo - Ensino Por CompetênciasDocument24 pagesArtigo - Ensino Por CompetênciasMarcelo Peixoto SouzaNo ratings yet
- Resenha - O ConceitoDocument2 pagesResenha - O ConceitoMarcelo Peixoto SouzaNo ratings yet
- Mini-Exame Do Estado Mental - MEEMDocument6 pagesMini-Exame Do Estado Mental - MEEMMarcelo Peixoto Souza100% (2)
- Iluminismo e sua contestaçãoDocument1 pageIluminismo e sua contestaçãoMarcelo Peixoto SouzaNo ratings yet
- Resumo - Texto Século XXDocument1 pageResumo - Texto Século XXMarcelo Peixoto SouzaNo ratings yet
- Fenomenologia em Michel HenryDocument10 pagesFenomenologia em Michel HenryMarcelo Peixoto SouzaNo ratings yet
- Relação Teoria e Prática Na Formação de ProfessoresDocument18 pagesRelação Teoria e Prática Na Formação de ProfessoresMarcelo Peixoto SouzaNo ratings yet
- Aop046 09Document8 pagesAop046 09Marcelo Peixoto SouzaNo ratings yet
- Treinamento de Habilidades Sociais Educativas para Pais de Crianças Com Problemas de ComportamentoDocument8 pagesTreinamento de Habilidades Sociais Educativas para Pais de Crianças Com Problemas de ComportamentoVictória NamurNo ratings yet
- Psicologia Geral I: Behaviorismo e HumanismoDocument7 pagesPsicologia Geral I: Behaviorismo e HumanismoHumbertoDomingosSérioNo ratings yet
- Relatório Do Estágio Supervisionado JINDocument17 pagesRelatório Do Estágio Supervisionado JINjeronimoinacio0% (1)
- O que é Cognição e seu SignificadoDocument6 pagesO que é Cognição e seu SignificadoIsolde KozowskiNo ratings yet
- Fichamento - Ocsana DanylukDocument2 pagesFichamento - Ocsana DanylukDrika HermioneNo ratings yet
- Esquizofrenia e psicoses brevesDocument3 pagesEsquizofrenia e psicoses brevesRafael MaltaNo ratings yet
- 8.simulado Tendências Pedagógicas - Docx 1Document10 pages8.simulado Tendências Pedagógicas - Docx 1Francis RamalhoNo ratings yet
- Filosofia da Linguagem: Curso EaD aborda pensadores e temas centraisDocument4 pagesFilosofia da Linguagem: Curso EaD aborda pensadores e temas centraismurilobsNo ratings yet
- Perspectivas Atuais Da Educação - GadottiDocument14 pagesPerspectivas Atuais Da Educação - GadottiHeloisa RibeiroNo ratings yet
- UNIP - Universidade Paulista - DisciplinaOnline - Sistemas de Conteúdo Online para Alunos 3Document15 pagesUNIP - Universidade Paulista - DisciplinaOnline - Sistemas de Conteúdo Online para Alunos 3MasomaNo ratings yet
- A Importância Da Metodologia Científica para Estudantes No Contexto UniversitárioDocument11 pagesA Importância Da Metodologia Científica para Estudantes No Contexto UniversitárioCrence AndreNo ratings yet
- A Aula como Unidade Estrutural do Processo de EnsinoDocument9 pagesA Aula como Unidade Estrutural do Processo de EnsinoarlindoNo ratings yet
- 1998 Beraldo D UfmtDocument190 pages1998 Beraldo D UfmtVinicius Machado Pereira dos santosNo ratings yet
- Meu Pé Esquerdo-RelatórioDocument8 pagesMeu Pé Esquerdo-RelatórioVictorNo ratings yet
- Aepal12 Ficha Form1Document9 pagesAepal12 Ficha Form1Isabel AraújoNo ratings yet
- Claudia GianettiDocument225 pagesClaudia GianettiLeonardo NajarNo ratings yet
- Sentimento de Pena: Contingências Complexas Das Quais É FunçãoDocument9 pagesSentimento de Pena: Contingências Complexas Das Quais É FunçãoJosé Hiroshi TanigutiNo ratings yet
- Curso ABA Autismo PetrolinaDocument9 pagesCurso ABA Autismo PetrolinaKarol Lala LalaNo ratings yet
- A importância do tradutorDocument6 pagesA importância do tradutorJerson BrennoNo ratings yet
- A ORDEM DO DISCURSO DE FOUCAULTDocument5 pagesA ORDEM DO DISCURSO DE FOUCAULTLara TeixeiraNo ratings yet
- Gestalt SlideDocument21 pagesGestalt SlideWanessa MatrizNo ratings yet
- Atas Cedu 13 Nas Pegadas Das Reformas Educativas PDFDocument470 pagesAtas Cedu 13 Nas Pegadas Das Reformas Educativas PDFEleutério AfonsoNo ratings yet
- Ficha de Dados Sobre Avaliação Do Aluno Surdo e Surdocego PDFDocument7 pagesFicha de Dados Sobre Avaliação Do Aluno Surdo e Surdocego PDFInstituto Helena Antipoff100% (3)
- Fatores e Processos de AprendizagemDocument26 pagesFatores e Processos de AprendizagemRita MonteiroNo ratings yet
- Guia - de - Estudos - de - Psicopedagogia NilvaDocument49 pagesGuia - de - Estudos - de - Psicopedagogia NilvaIrisnei NunesNo ratings yet
- 619-Texto Do Artigo-2031-2220-10-20230429Document16 pages619-Texto Do Artigo-2031-2220-10-20230429Aline EngelNo ratings yet
- APEL'S PHILOSOPHY AND ITS IMPACT ON LAWDocument20 pagesAPEL'S PHILOSOPHY AND ITS IMPACT ON LAWLucas Chagas100% (1)
- Estratégias de Leitura - Ismar Souza PDFDocument80 pagesEstratégias de Leitura - Ismar Souza PDFTito PachecoNo ratings yet
- Evolução da língua portuguesa no BrasilDocument16 pagesEvolução da língua portuguesa no BrasilValter Mário Pereira JúniorNo ratings yet