You might also like
- Url-2003 Parvovirus en Neuronas de GatoDocument5 pagesUrl-2003 Parvovirus en Neuronas de GatocristianNo ratings yet
- 104063870001200209Document3 pages104063870001200209Kent Solis VicenteNo ratings yet
- Apoptosis in Natural Rabies Virus Infection in DogsDocument5 pagesApoptosis in Natural Rabies Virus Infection in DogspapiipiiNo ratings yet
- Feline Panleukopenia Virus Infection in A Captive-Bred Bengal Tiger (AND A LeopardDocument4 pagesFeline Panleukopenia Virus Infection in A Captive-Bred Bengal Tiger (AND A LeopardzulfikarNo ratings yet
- EchinococcusDocument20 pagesEchinococcusJuan Fernando Calcina IsiqueNo ratings yet
- Occurrence of Ferret Enteric Coronavirus in Brazil (Preliminary Report)Document3 pagesOccurrence of Ferret Enteric Coronavirus in Brazil (Preliminary Report)Yovan BlancoNo ratings yet
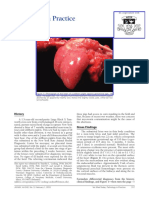
- Patología en La PrácticaDocument3 pagesPatología en La PrácticaSTAN BLACKNo ratings yet
- Characterization and Complete Genome Sequence of A Novel Coronavirus, Coronavirus HKU1, From Patients With PneumoniaDocument12 pagesCharacterization and Complete Genome Sequence of A Novel Coronavirus, Coronavirus HKU1, From Patients With PneumoniaAlexandra PaunNo ratings yet
- Helicobacter Pylori: Upregulates Prion Protein Expression in Gastric Mucosa: A Possible Link To Prion DiseaseDocument6 pagesHelicobacter Pylori: Upregulates Prion Protein Expression in Gastric Mucosa: A Possible Link To Prion DiseaseNadeem MalikNo ratings yet
- Articulo 1 InmunogenomicaDocument6 pagesArticulo 1 InmunogenomicaMichelle PoulainNo ratings yet
- Dog with Acute Hepatozoon canis InfectionDocument4 pagesDog with Acute Hepatozoon canis InfectionSivapong VsmuNo ratings yet
- What Is Your Diagnosis? Blood Smear From A Cat: Mary Leissinger, Stuart Walton, Jon Fletcher, Stephen GauntDocument2 pagesWhat Is Your Diagnosis? Blood Smear From A Cat: Mary Leissinger, Stuart Walton, Jon Fletcher, Stephen GauntGharbi KamiliaNo ratings yet
- Allele Frequency of Nonsense Mutation Responsible For He - 2020 - Veterinary andDocument3 pagesAllele Frequency of Nonsense Mutation Responsible For He - 2020 - Veterinary andangelthaiboxyahoo.esNo ratings yet
- Evaluation of PDNS induction in gnotobiotic pigs rules out PCV2Document8 pagesEvaluation of PDNS induction in gnotobiotic pigs rules out PCV2Matthew HsuNo ratings yet
- Canine Coronavirus-Associated Puppy Mortality Without Evidence of Concurrent Canine Parvovirus InfectionDocument5 pagesCanine Coronavirus-Associated Puppy Mortality Without Evidence of Concurrent Canine Parvovirus Infectionhein heinNo ratings yet
- Progression of Pulmonary Lesions in Cat with ParagonimosisDocument5 pagesProgression of Pulmonary Lesions in Cat with ParagonimosisMahdawwNo ratings yet
- Adapted Tembusu-Like Virus in Chickens and Geese in ChinaDocument3 pagesAdapted Tembusu-Like Virus in Chickens and Geese in ChinaTrung Kien LeNo ratings yet
- Immunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient MouseDocument5 pagesImmunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient Mouselily rogaNo ratings yet
- Kutasi Et Al-2011-Journal of Veterinary Internal Medicine PDFDocument6 pagesKutasi Et Al-2011-Journal of Veterinary Internal Medicine PDFAnonymous uCvzE0NVNo ratings yet
- CVJ 10 1091Document3 pagesCVJ 10 1091Rayza LubisNo ratings yet
- B The PigDocument16 pagesB The PigMaria SerlyantiNo ratings yet
- Concentraciones Sericas de Eritropoyetina en Perro y Gatos Anemicos y PolicitemicosDocument8 pagesConcentraciones Sericas de Eritropoyetina en Perro y Gatos Anemicos y PolicitemicosZoleida BastidasNo ratings yet
- Sanchez Cordon Et Al 2005 Lymphocyte Apoptosis and Thrombocytopenia in Spleen During Classical Swine Fever Role OfDocument12 pagesSanchez Cordon Et Al 2005 Lymphocyte Apoptosis and Thrombocytopenia in Spleen During Classical Swine Fever Role OfKeto PrehranaNo ratings yet
- 2002-Specific Bovine Antibody Response Against A New Recombinant Cryptosporidium Parvum Antigen Containing 4 Zinc-Finger MotifsDocument6 pages2002-Specific Bovine Antibody Response Against A New Recombinant Cryptosporidium Parvum Antigen Containing 4 Zinc-Finger MotifswiwienNo ratings yet
- BF01309668Document13 pagesBF01309668pafspontesNo ratings yet
- Nothing To DoDocument11 pagesNothing To DoDiana Muela MoraNo ratings yet
- Detection NHP Texa and PeruDocument6 pagesDetection NHP Texa and PerurenatogouveiapNo ratings yet
- Jurnal 1 KalsitoninDocument9 pagesJurnal 1 KalsitoninPrawido AndriadiNo ratings yet
- JM 002076Document5 pagesJM 002076Malú CarranzaNo ratings yet
- Alleman (1999) - Western Immunoblot Analysis of The Antigens ofDocument6 pagesAlleman (1999) - Western Immunoblot Analysis of The Antigens ofWalison LoiolaNo ratings yet
- Encefalopatia Uremica EquinaDocument5 pagesEncefalopatia Uremica EquinaDavid CuervoNo ratings yet
- Acute Opisthorchiasis in A Kazakh ManDocument2 pagesAcute Opisthorchiasis in A Kazakh Manhusna aninda farahitaNo ratings yet
- Advpub 16-0463 PDFDocument20 pagesAdvpub 16-0463 PDFMarianaNo ratings yet
- Deteksi Virus Classical Swine Fever Di Bali Dengan RT-PCR PDFDocument8 pagesDeteksi Virus Classical Swine Fever Di Bali Dengan RT-PCR PDFMuhammadIbrahimNo ratings yet
- Virulence Factors, Antibiotic Resistance Phenotypes and O-Serogroups of Escherichia Coli Strains Isolated From Community-Acquired Urinary Tract Infection Patients in MexicoDocument8 pagesVirulence Factors, Antibiotic Resistance Phenotypes and O-Serogroups of Escherichia Coli Strains Isolated From Community-Acquired Urinary Tract Infection Patients in MexicounicarpNo ratings yet
- The Veterinary JournalDocument6 pagesThe Veterinary JournalMustafa ERSOYNo ratings yet
- Plasmodium Knowlesi: Human Infection Detected by Rapid Diagnostic Tests For MalariaDocument3 pagesPlasmodium Knowlesi: Human Infection Detected by Rapid Diagnostic Tests For MalariaAlvin PratamaNo ratings yet
- Esc Pointers On The Diagnosis and Administration of Acute Pulmonary EmbolismDocument3 pagesEsc Pointers On The Diagnosis and Administration of Acute Pulmonary EmbolismgwaniegdnrNo ratings yet
- Buhles 1974Document12 pagesBuhles 1974Paula Andrea Castro MartinezNo ratings yet
- Vet Pathol 2011 2011 ACVP Annual Meeting E1 E51Document52 pagesVet Pathol 2011 2011 ACVP Annual Meeting E1 E51Haroon RashidNo ratings yet
- Feline Gastrointestinal Eosinophilic Sclerosing FibroplasiaDocument8 pagesFeline Gastrointestinal Eosinophilic Sclerosing FibroplasiaCarlos Alberto Chaves VelasquezNo ratings yet
- Psittacid Herpesvirus 3 Infection in The Eclectus Parrot (Eclectus Roratus) in AustraliaDocument5 pagesPsittacid Herpesvirus 3 Infection in The Eclectus Parrot (Eclectus Roratus) in AustraliaJeann LealNo ratings yet
- Https:journals Sagepub Com:doi:pdf:10 1354:vp 37-6-650Document3 pagesHttps:journals Sagepub Com:doi:pdf:10 1354:vp 37-6-650Lucas XavierNo ratings yet
- Veterinary Internal Medicne - 2008 - Hartmann - Comparison of Different Tests To Diagnose Feline Infectious PeritonitisDocument10 pagesVeterinary Internal Medicne - 2008 - Hartmann - Comparison of Different Tests To Diagnose Feline Infectious Peritonitisxn5q8nckvkNo ratings yet
- ARTICLE REMO Bacterial PericarditisDocument3 pagesARTICLE REMO Bacterial PericarditisDulNo ratings yet
- Barrerameja2009 2Document6 pagesBarrerameja2009 2erzebeth bathoryNo ratings yet
- Detection of Yersinia Pestis in Sputum by Real-Time PCRDocument3 pagesDetection of Yersinia Pestis in Sputum by Real-Time PCRverNo ratings yet
- Prevalence of Enterotoxigenic Clostridium Perfringens in Meats in San Luis, ArgentinaDocument6 pagesPrevalence of Enterotoxigenic Clostridium Perfringens in Meats in San Luis, Argentinamauro manfrediNo ratings yet
- Benetka2006 Canine Adenovirus Type 2 Infection in Four Puppies With Neurological SignsDocument6 pagesBenetka2006 Canine Adenovirus Type 2 Infection in Four Puppies With Neurological SignsMaría JoséNo ratings yet
- Pathogenesis Canine Parvovirus Enteritis: The Importance of ViremiaDocument12 pagesPathogenesis Canine Parvovirus Enteritis: The Importance of ViremiaDiana OntiverosNo ratings yet
- Tian 2016Document13 pagesTian 2016Epiphany SonderNo ratings yet
- The Sites of Early Viral Replication in FelineDocument13 pagesThe Sites of Early Viral Replication in FelineMinaduki KashouNo ratings yet
- Octogon DegusDocument6 pagesOctogon DegusKaren Esteves ZúñigaNo ratings yet
- Persistence of Porcine Reproductive and Virus in Intensive Farrow-to-Finish Pig HerdsDocument7 pagesPersistence of Porcine Reproductive and Virus in Intensive Farrow-to-Finish Pig HerdsJuan Fernando Calcina IsiqueNo ratings yet
- Effect of Antibodies On The Expression of Plasmodium Falciparum Circumsporozoite Protein GeneDocument4 pagesEffect of Antibodies On The Expression of Plasmodium Falciparum Circumsporozoite Protein Genereza gomaNo ratings yet
- Anaplasma Phagocytophilum, Bartonella SPP., Haemoplasma Species and Hepatozoon Spp. in Ticks Infesting Cats: A Large-Scale SurveyDocument10 pagesAnaplasma Phagocytophilum, Bartonella SPP., Haemoplasma Species and Hepatozoon Spp. in Ticks Infesting Cats: A Large-Scale SurveyValentina RondonNo ratings yet
- Pagination VETPAR 109611Document13 pagesPagination VETPAR 109611Bárbara Laurice Araújo VerçosaNo ratings yet
- New Viruses in Idiopathic Human Diarrhea Cases, The NetherlandsDocument5 pagesNew Viruses in Idiopathic Human Diarrhea Cases, The NetherlandsMark ReinhardtNo ratings yet
- Evidence For Transmission of Plasmodium Vivax Among A Duffy Antigen Negative Population in Western KenyaDocument7 pagesEvidence For Transmission of Plasmodium Vivax Among A Duffy Antigen Negative Population in Western Kenyamaugui18No ratings yet
- Pathophysiology of Parasitic InfectionFrom EverandPathophysiology of Parasitic InfectionE SoulsbyNo ratings yet