You might also like
- Pretreatment and hydrolysis of cellulosic agricultural wastes for bioethanol productionDocument7 pagesPretreatment and hydrolysis of cellulosic agricultural wastes for bioethanol productionAndré Felipe SilvaNo ratings yet
- 14043-Article Text PDF-58576-2-10-20190902Document8 pages14043-Article Text PDF-58576-2-10-20190902R. VersteeghNo ratings yet
- ABAB 2006 SeparationDocument13 pagesABAB 2006 SeparationJulieth Cardona C.No ratings yet
- Membranes: Continuous Ethanol Production With A Membrane Bioreactor at High Acetic Acid ConcentrationsDocument16 pagesMembranes: Continuous Ethanol Production With A Membrane Bioreactor at High Acetic Acid ConcentrationsR-A PascualNo ratings yet
- Kinetic Simple GoodDocument11 pagesKinetic Simple GoodReza MahoorNo ratings yet
- Process Intensification in Lactic Acid ProductionDocument11 pagesProcess Intensification in Lactic Acid ProductionRoxana StoicaNo ratings yet
- Acido Lactico Hojarasca de MaizDocument34 pagesAcido Lactico Hojarasca de MaizAlex CárdenasNo ratings yet
- Lactosuero 9 NoDocument24 pagesLactosuero 9 NoMarco Antonio Salcedo HinojosaNo ratings yet
- J Biortech 2017 12 033Document8 pagesJ Biortech 2017 12 033fajarNo ratings yet
- Silva 2012Document7 pagesSilva 2012Brian Oro BeltránNo ratings yet
- Journal Pone 0107143Document6 pagesJournal Pone 0107143suryasanNo ratings yet
- J Biombioe 2010 02 003Document7 pagesJ Biombioe 2010 02 003ABDUL ZIRAY ARIFINNo ratings yet
- Oonkhanond 2017Document37 pagesOonkhanond 2017Gebby FebriliaNo ratings yet
- Fermentation and Crystallization of Succinic Acid From Actinobacillus Succinogenes ATCC55618 Using Fresh Cassava Root As The Main SubstrateDocument11 pagesFermentation and Crystallization of Succinic Acid From Actinobacillus Succinogenes ATCC55618 Using Fresh Cassava Root As The Main SubstrateYudid Anaya CaroNo ratings yet
- Optimization of Chemical Pretreatments Using Response Surface Methodology For Second-Generation Ethanol Production From Coffee Husk WasteDocument13 pagesOptimization of Chemical Pretreatments Using Response Surface Methodology For Second-Generation Ethanol Production From Coffee Husk WasteAngie Estefania Rivera CordobaNo ratings yet
- Artículo PDFDocument7 pagesArtículo PDFEric Adiel Lona GaribayNo ratings yet
- Production of Ethanol From Soybean Hull Hydrolysate by Osmotolerant Candida Guilliermondii NRRL Y-2075Document7 pagesProduction of Ethanol From Soybean Hull Hydrolysate by Osmotolerant Candida Guilliermondii NRRL Y-2075Camila NevesNo ratings yet
- Omay 2012Document8 pagesOmay 201215041168No ratings yet
- Separation and Purification TechnologyDocument7 pagesSeparation and Purification TechnologyIka SulistyaningtiyasNo ratings yet
- Potential and Assessment of Lactic Acid Production and Isolation - A ReviewDocument34 pagesPotential and Assessment of Lactic Acid Production and Isolation - A ReviewRenata CamilaNo ratings yet
- Yi-Huang Chang, Ku-Shang Chang, Cheng-Wei Huang, Chuan-Liang Hsu, Hung-Der JangDocument8 pagesYi-Huang Chang, Ku-Shang Chang, Cheng-Wei Huang, Chuan-Liang Hsu, Hung-Der JangNoor SalamNo ratings yet
- Pretreatment Conditions of Rice Straw For Simultaneous Hydrogen and Ethanol Fermentation by Mixed CultureDocument8 pagesPretreatment Conditions of Rice Straw For Simultaneous Hydrogen and Ethanol Fermentation by Mixed CultureAldrienNo ratings yet
- Industrial Crops and Products: Talita M. Lacerda, Márcia D. Zambon, Elisabete FrolliniDocument10 pagesIndustrial Crops and Products: Talita M. Lacerda, Márcia D. Zambon, Elisabete FrolliniMuhammadFakhrizalFahmiNo ratings yet
- 2cost-Effective Simultaneous Saccharification and Fermentation of L-LacticDocument8 pages2cost-Effective Simultaneous Saccharification and Fermentation of L-LacticOlga Lucia BayonaNo ratings yet
- Integrated Production of Second Generation Ethanol and Lactic Acid From Steam-Exploded Elephant GrassDocument8 pagesIntegrated Production of Second Generation Ethanol and Lactic Acid From Steam-Exploded Elephant Grassluana.rochaNo ratings yet
- A Novel Cell Factory For Efficient Production of Ethanol From Dairy WasteDocument11 pagesA Novel Cell Factory For Efficient Production of Ethanol From Dairy WasteVitor ArsenioNo ratings yet
- Pilot-Scale Lactic Acid Production Via Batch Culturing of Lactobacillus Sp. RKY2 Using Corn Steep Liquor As A Nitrogen SourceDocument6 pagesPilot-Scale Lactic Acid Production Via Batch Culturing of Lactobacillus Sp. RKY2 Using Corn Steep Liquor As A Nitrogen SourcekopisusumantapNo ratings yet
- Fermentative lactic acid production from coffee pulpDocument7 pagesFermentative lactic acid production from coffee pulpGebby FebriliaNo ratings yet
- Fermentation of Lactose To Ethanol in Cheese WheyDocument13 pagesFermentation of Lactose To Ethanol in Cheese WheyVitor ArsenioNo ratings yet
- Effect of Dilute Acid Pretreatment of Different Energy Crops On Their Enzymatic HydrolysisDocument13 pagesEffect of Dilute Acid Pretreatment of Different Energy Crops On Their Enzymatic HydrolysisProAkademiaNo ratings yet
- Enzymatic Saccharification and Fermentation of CelDocument10 pagesEnzymatic Saccharification and Fermentation of CelSharmaNo ratings yet
- Waste ManagementDocument11 pagesWaste ManagementTidar MelianiNo ratings yet
- Chemical Conversion of Biomass To Green ChemicalsDocument32 pagesChemical Conversion of Biomass To Green ChemicalsMaria Cecille Sarmiento GarciaNo ratings yet
- Artículo Ácido LacticoDocument7 pagesArtículo Ácido LacticoXavier Castro CentenoNo ratings yet
- Liu 2015Document8 pagesLiu 2015Renata CamilaNo ratings yet
- Optimization of soybean hull acid hydrolysis for bioconversionDocument9 pagesOptimization of soybean hull acid hydrolysis for bioconversionCamila NevesNo ratings yet
- Acido SuccinicoDocument17 pagesAcido SuccinicoLuisa Gomez RomanNo ratings yet
- General Characteristics and Treatment Possibilities of Dairy Wastewater - A ReviewDocument15 pagesGeneral Characteristics and Treatment Possibilities of Dairy Wastewater - A Reviewangelapaul peterNo ratings yet
- Biocatalysis and Agricultural Biotechnology: SciencedirectDocument8 pagesBiocatalysis and Agricultural Biotechnology: SciencedirectAyush SinghNo ratings yet
- Rice FtirDocument15 pagesRice FtirRavindra AgarwalNo ratings yet
- A Review On The Current Developments in Continuous Lactic Acid Fermentations and Case Studies Utilising Inexpensive Raw MaterialsDocument10 pagesA Review On The Current Developments in Continuous Lactic Acid Fermentations and Case Studies Utilising Inexpensive Raw MaterialsIntanFauziahNo ratings yet
- Improving Biogas Production From Wheat Plant Using Alkaline PretreatmentDocument6 pagesImproving Biogas Production From Wheat Plant Using Alkaline PretreatmentMaryam TaherdanakNo ratings yet
- Pretreatment and Saccharification of Rice HuksDocument7 pagesPretreatment and Saccharification of Rice HuksHimpunan Mahasiswa Teknik KimiaNo ratings yet
- Cellulolysis - Ethanol Production: Cellulolysis Is The Process of Breaking Down Cellulose IntoDocument9 pagesCellulolysis - Ethanol Production: Cellulolysis Is The Process of Breaking Down Cellulose Intoanushri birlaNo ratings yet
- Fermentation of Lactose To Ethanol in Cheese Whey Permeate and Concentrated Permeate by Engineered Escherichia ColiDocument12 pagesFermentation of Lactose To Ethanol in Cheese Whey Permeate and Concentrated Permeate by Engineered Escherichia ColijoelfloresanchezNo ratings yet
- Synthesis and Biological Application of PolylacticDocument18 pagesSynthesis and Biological Application of PolylacticNelson Enrique Bessone MadridNo ratings yet
- C. Cohnii Fed-BatchDocument7 pagesC. Cohnii Fed-BatchLucilaFernandezNo ratings yet
- 2010 AndreanneHarbecDocument189 pages2010 AndreanneHarbecSjjshahaNo ratings yet
- Acid Hydrolysis of Sugarcane Bagasse For Lactic Acid ProductionDocument8 pagesAcid Hydrolysis of Sugarcane Bagasse For Lactic Acid Productiondh2vuihocNo ratings yet
- Enhanced Lactic Acid Production From Carob Extract by Lactobacillus Casei Using Invertase Pretreatment PDFDocument12 pagesEnhanced Lactic Acid Production From Carob Extract by Lactobacillus Casei Using Invertase Pretreatment PDFRafael RubioNo ratings yet
- Solowiej2016 - Processed Cheese AnaloguesDocument9 pagesSolowiej2016 - Processed Cheese AnaloguesDinoNo ratings yet
- Brazilian Journal of Chemical Engineering - Fermentable sugars from water hyacinthDocument9 pagesBrazilian Journal of Chemical Engineering - Fermentable sugars from water hyacinthJerome JavierNo ratings yet
- 14bdo - Pdfsuccinic Acid: A New Platform Chemical For Biobased Polymers From Renewable ResourcesDocument8 pages14bdo - Pdfsuccinic Acid: A New Platform Chemical For Biobased Polymers From Renewable ResourcesAngela Smith100% (1)
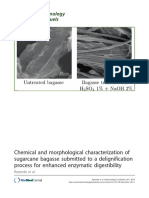
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- ViewFreeArticle PDFDocument5 pagesViewFreeArticle PDFMaria Alyssa EdañoNo ratings yet
- Research Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionDocument10 pagesResearch Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionShankar AcharNo ratings yet
- Evaluation of Support Matrices For Immobilization of Anaerobic Consortia For e Cient Carbon Cycling in Waste RegenerationDocument10 pagesEvaluation of Support Matrices For Immobilization of Anaerobic Consortia For e Cient Carbon Cycling in Waste RegenerationGregorius BudiantoNo ratings yet
- Lactic Acid From Apple Pomace A Laboratory Experiment For Teaching Valorisation of Wastes-ExpDocument7 pagesLactic Acid From Apple Pomace A Laboratory Experiment For Teaching Valorisation of Wastes-ExpFizza MaryamNo ratings yet
- Polar Lipids: Biology, Chemistry, and TechnologyFrom EverandPolar Lipids: Biology, Chemistry, and TechnologyMoghis U. AhmadNo ratings yet
- Plant and Animal Bio-Chemistry - Including Information on Amino Acids, Proteins, Pigments and Other Chemical Constituents of Organic MatterFrom EverandPlant and Animal Bio-Chemistry - Including Information on Amino Acids, Proteins, Pigments and Other Chemical Constituents of Organic MatterNo ratings yet
- Bioactive Compounds in Potatoes: Accumulation Under Drought Stress ConditionsDocument9 pagesBioactive Compounds in Potatoes: Accumulation Under Drought Stress ConditionsComan GigiNo ratings yet
- Obtinere Acid Citric Din Coji de Portocale Utilizand A.niger PDFDocument16 pagesObtinere Acid Citric Din Coji de Portocale Utilizand A.niger PDFComan GigiNo ratings yet
- Producția Etanolului Continuă de Kluyveromyces SP PDFDocument6 pagesProducția Etanolului Continuă de Kluyveromyces SP PDFComan GigiNo ratings yet
- Article 9 Fatma PDFDocument8 pagesArticle 9 Fatma PDFComan GigiNo ratings yet
- Jce2015 PDFDocument9 pagesJce2015 PDFComan GigiNo ratings yet
- Castro-Gómez, 2014 (A High Performance Transmethylation Protocol)Document6 pagesCastro-Gómez, 2014 (A High Performance Transmethylation Protocol)Coman GigiNo ratings yet
- Characterization of Alkaliphilic Surfactant Stable and Raw Starch Digesting Amylase From Bacillus Subtilis Strain Js 1948 5948.S8 002 PDFDocument6 pagesCharacterization of Alkaliphilic Surfactant Stable and Raw Starch Digesting Amylase From Bacillus Subtilis Strain Js 1948 5948.S8 002 PDFComan GigiNo ratings yet
- Bioactive Compounds in Potatoes: Accumulation Under Drought Stress ConditionsDocument9 pagesBioactive Compounds in Potatoes: Accumulation Under Drought Stress ConditionsComan GigiNo ratings yet
- Yea0031 0127 PDFDocument9 pagesYea0031 0127 PDFComan GigiNo ratings yet
- Alginate PDFDocument74 pagesAlginate PDFComan GigiNo ratings yet
- Aplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFDocument10 pagesAplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFComan GigiNo ratings yet
- Aplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFDocument10 pagesAplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFComan GigiNo ratings yet
- Patheon Am 14 1836 Lyophilisation Parameters PDFDocument4 pagesPatheon Am 14 1836 Lyophilisation Parameters PDFComan GigiNo ratings yet
- 2 PDFDocument10 pages2 PDFComan GigiNo ratings yet
- Ribeiro412013BBJ6066 1 PDFDocument10 pagesRibeiro412013BBJ6066 1 PDFComan GigiNo ratings yet
- Alginate PDFDocument74 pagesAlginate PDFComan GigiNo ratings yet
- Bioactive Compounds in Potatoes: Accumulation Under Drought Stress ConditionsDocument9 pagesBioactive Compounds in Potatoes: Accumulation Under Drought Stress ConditionsComan GigiNo ratings yet
- Haba Et Al IJP2014 PDFDocument8 pagesHaba Et Al IJP2014 PDFComan GigiNo ratings yet
- Melguiribacillus Thermohalophilus PDFDocument10 pagesMelguiribacillus Thermohalophilus PDFComan GigiNo ratings yet
- Aplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFDocument10 pagesAplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFComan GigiNo ratings yet
- Aplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFDocument10 pagesAplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFComan GigiNo ratings yet
- Article Caldicobacter 2014 PDFDocument13 pagesArticle Caldicobacter 2014 PDFComan GigiNo ratings yet
- Castro-Gómez, 2014 (A High Performance Transmethylation Protocol)Document6 pagesCastro-Gómez, 2014 (A High Performance Transmethylation Protocol)Coman GigiNo ratings yet
- Fmicb 05 00708 PDFDocument15 pagesFmicb 05 00708 PDFComan GigiNo ratings yet
- An Integrative Process For Bio-Ethanol Production Employing SSF Produced Cellulase Without Extraction PDFDocument12 pagesAn Integrative Process For Bio-Ethanol Production Employing SSF Produced Cellulase Without Extraction PDFComan GigiNo ratings yet
- 2 PDFDocument10 pages2 PDFComan GigiNo ratings yet
- Article 9 Fatma PDFDocument8 pagesArticle 9 Fatma PDFComan GigiNo ratings yet
- Aplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFDocument10 pagesAplicarea de Micronutrienti Pe Baza de Calciu Si Molibden Foliar PDFComan GigiNo ratings yet
- 5419-24327-1-PB Bioresorce PDFDocument36 pages5419-24327-1-PB Bioresorce PDFComan GigiNo ratings yet
- Aiba 1968 Kinetics of Product Inhibition in Alcohol FermentationDocument20 pagesAiba 1968 Kinetics of Product Inhibition in Alcohol Fermentationrishu2525No ratings yet
- Seminars in Orthodontics 2005 CarlsonDocument12 pagesSeminars in Orthodontics 2005 CarlsonMariana Levy100% (1)
- Cerca de 1,910,000 Resultados (0.26 Segundos) : RGD - Mcw.eduDocument2 pagesCerca de 1,910,000 Resultados (0.26 Segundos) : RGD - Mcw.edutlowdidj33No ratings yet
- pGEM-T EasyDocument15 pagespGEM-T EasyRocío Romero HinojosaNo ratings yet
- AlleleBalanced Luciferase Assay KitsDocument1 pageAlleleBalanced Luciferase Assay KitsAlleleBiotechNo ratings yet
- NBlotDocument11 pagesNBlotmaulidyaNo ratings yet
- Single Cell Models, Shuler 1999Document4 pagesSingle Cell Models, Shuler 1999Marcelo Martinez CajigasNo ratings yet
- Universal Haplotype-Based Noninvasive Prenatal Testing For Single Gene DiseasesDocument12 pagesUniversal Haplotype-Based Noninvasive Prenatal Testing For Single Gene DiseasestestNo ratings yet
- Metabolomics BasicsDocument37 pagesMetabolomics Basicsvijay2109No ratings yet
- Genetic Heritage of Croatians in The Southeastern European Gene Pool-Y Chromosome Analysis of The Croatian Continental and Island PopulationDocument9 pagesGenetic Heritage of Croatians in The Southeastern European Gene Pool-Y Chromosome Analysis of The Croatian Continental and Island PopulationJan KowalskiNo ratings yet
- Bread Making Techniques: Traditional vs Modern MethodsDocument8 pagesBread Making Techniques: Traditional vs Modern MethodsSyraJae24No ratings yet
- INFORS HT White Paper Continuous CultureDocument6 pagesINFORS HT White Paper Continuous CulturebiocloneNo ratings yet
- Gmo Crops-Research PaperDocument15 pagesGmo Crops-Research Paperapi-381986176No ratings yet
- Role of Biotechnology in Improving Human HealthDocument7 pagesRole of Biotechnology in Improving Human HealthdeepaksagarNo ratings yet
- Biology Lab 1: Bioinformatic Report AnalysisDocument5 pagesBiology Lab 1: Bioinformatic Report AnalysisKasia DrewniakNo ratings yet
- Indian PharmaDocument32 pagesIndian PharmaNidhiNo ratings yet
- NCS TCP GuidelinesDocument32 pagesNCS TCP GuidelinesSandhya SinghNo ratings yet
- Differences Between Mitosis and MeiosisDocument4 pagesDifferences Between Mitosis and MeiosisMeri SunderNo ratings yet
- PCR Master Mix ProtocolDocument2 pagesPCR Master Mix ProtocolDo Thi Van AnhNo ratings yet
- CHO HCP Poster Presented at Bangalore India Bio2011Document1 pageCHO HCP Poster Presented at Bangalore India Bio2011mercerexpNo ratings yet
- Genetic Diversity of Total, Active and Culturable Marine Bacteria in Coastal SeawaterDocument11 pagesGenetic Diversity of Total, Active and Culturable Marine Bacteria in Coastal SeawaterMelody ChristineNo ratings yet
- SAFC Biosciences - Technical Bulletin - EX-CELL™ CD CHO: A Chemically Defined, Animal-Component Free, Serum-Free Medium For CHO CellsDocument4 pagesSAFC Biosciences - Technical Bulletin - EX-CELL™ CD CHO: A Chemically Defined, Animal-Component Free, Serum-Free Medium For CHO CellsSAFC-GlobalNo ratings yet
- Audit Penggunaan Aantibiotik Secara Kuantitatif & Kualitatif PDFDocument9 pagesAudit Penggunaan Aantibiotik Secara Kuantitatif & Kualitatif PDFabd. rahim mubarakNo ratings yet
- 17-10018 T-SeriesTFF Cassettes SS USD2322c SSDocument4 pages17-10018 T-SeriesTFF Cassettes SS USD2322c SSCHIRANJEEVINo ratings yet
- New Bioprocessing Strategies: Development and Manufacturing of Recombinant Antibodies and ProteinsDocument473 pagesNew Bioprocessing Strategies: Development and Manufacturing of Recombinant Antibodies and Proteinsmohammad100% (1)
- Composición Extracto de LevaduraDocument1 pageComposición Extracto de LevaduraJuank González TéllezNo ratings yet
- Centogene Indonesia CME 190830Document49 pagesCentogene Indonesia CME 190830Arshiya Nayara AvanishaNo ratings yet
- Bio Drosophilia Mini Project 2 1Document15 pagesBio Drosophilia Mini Project 2 1jocyeo0% (1)
- Gest 4 21 2010Document228 pagesGest 4 21 2010optimusnowNo ratings yet
- Techniques in Molecular Biology and Dna TechnologyDocument104 pagesTechniques in Molecular Biology and Dna TechnologyMd Shahroz AlamNo ratings yet