You might also like
- Challenging the Limits of the Human Life Span: - Can We Live Longer Than 120 Years - New GuidelinesFrom EverandChallenging the Limits of the Human Life Span: - Can We Live Longer Than 120 Years - New GuidelinesNo ratings yet
- Dr. Bernstein's Diabetes Solution - The Complete Guide To Achieving Normal Blood Sugars (PDFDrive) - 3Document10 pagesDr. Bernstein's Diabetes Solution - The Complete Guide To Achieving Normal Blood Sugars (PDFDrive) - 3ArtackNo ratings yet
- Human Mineral 2Document24 pagesHuman Mineral 2Nitay SorkarNo ratings yet
- Antiaging 101: Course Manual: A Proactive Preventive Health Care ProgramFrom EverandAntiaging 101: Course Manual: A Proactive Preventive Health Care ProgramNo ratings yet
- Digestive ImbalancesDocument73 pagesDigestive ImbalancesMuhammad Ricky Julian AdhetiaNo ratings yet
- Gonad Body Type Eating PlanDocument4 pagesGonad Body Type Eating PlanRosa María Rodríguez de PaoliNo ratings yet
- Miracles of Islam - DNA Genetic EngineeringDocument29 pagesMiracles of Islam - DNA Genetic Engineeringahmad kajNo ratings yet
- Diabetes Management: OverviewDocument20 pagesDiabetes Management: Overviewlaloo01No ratings yet
- How Is Diabetes Managed?: Type 1 and Type 2 Diabetes ORDocument2 pagesHow Is Diabetes Managed?: Type 1 and Type 2 Diabetes ORdebabrata5976No ratings yet
- Topical Corticosteroid AbuseDocument34 pagesTopical Corticosteroid AbuseDr Daulat Ram DhakedNo ratings yet
- Diabetes InformationDocument171 pagesDiabetes Informationtheia28No ratings yet
- How Do I Cleanse Myself SuccessfullyDocument8 pagesHow Do I Cleanse Myself SuccessfullyTony TshouNo ratings yet
- Truth About DiabetesDocument12 pagesTruth About DiabetesAmyHolderWilsonNo ratings yet
- 180 Degree DiabetesDocument111 pages180 Degree DiabetesJoRa71No ratings yet
- How To Use Amino Acids Like To End Sugar Cravings and Ease AnxietyDocument8 pagesHow To Use Amino Acids Like To End Sugar Cravings and Ease AnxietySK100% (1)
- Copper DeficiencyDocument30 pagesCopper DeficiencyMathewos MekonnenNo ratings yet
- The Prime Cause and Prevention of Disease WarburgDocument3 pagesThe Prime Cause and Prevention of Disease WarburgMichael DietrickNo ratings yet
- Surprising Reason Why You May Need More Carbs PDFDocument14 pagesSurprising Reason Why You May Need More Carbs PDFrazvan rotaru100% (1)
- Pathophysiology Cellulite and Treatment 2nd Edition Basic and Clinical DermatologyDocument19 pagesPathophysiology Cellulite and Treatment 2nd Edition Basic and Clinical DermatologySri Nauli Dewi LubisNo ratings yet
- Insulin Resistance & Green TeaDocument6 pagesInsulin Resistance & Green TeaSrinivasan GanapathiNo ratings yet
- 3 BIOHACKS For Energy Longevity Report 2023 10 10Document69 pages3 BIOHACKS For Energy Longevity Report 2023 10 10jahiddleNo ratings yet
- Smart Blood SugarDocument3 pagesSmart Blood SugarSimon Josh0% (1)
- NUTRILITE ProteinPowder en UsDocument5 pagesNUTRILITE ProteinPowder en UsFauzan IbrahimNo ratings yet
- Diabetes Millitus PDFDocument41 pagesDiabetes Millitus PDFAbdullah BhattiNo ratings yet
- Caffeine: The Good, The Bad, and The UglyDocument4 pagesCaffeine: The Good, The Bad, and The UglyBerenice RomeroNo ratings yet
- Start Low and Go Slow 1-21-2014Document5 pagesStart Low and Go Slow 1-21-2014Ed JonesNo ratings yet
- Ihcanapril17 PDFDocument48 pagesIhcanapril17 PDFMommyx44100% (2)
- Brain Exercises That Boost Memory PDFDocument3 pagesBrain Exercises That Boost Memory PDFPradeep KumarNo ratings yet
- Your Brain at Work: The Big IdeasDocument6 pagesYour Brain at Work: The Big IdeasInfinit13No ratings yet
- Health Seminar PresentationDocument46 pagesHealth Seminar Presentationmr_harshahsNo ratings yet
- Cell Phone Radiation - 85% Eliminated With Military Stealth TechnologyDocument8 pagesCell Phone Radiation - 85% Eliminated With Military Stealth TechnologyJohn DavidNo ratings yet
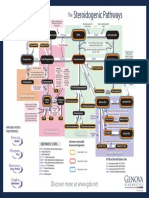
- The Steroidogenic PathwaysDocument1 pageThe Steroidogenic PathwaysMarkus van der WesthuizenNo ratings yet
- Energetic Intervention Update - 03 - March - 2018Document14 pagesEnergetic Intervention Update - 03 - March - 2018Maela Paliaga Budim0% (1)
- Chris Masterjohn Book DemoDocument15 pagesChris Masterjohn Book Demojijibiji24100% (1)
- Health Implications of A Vegetarian Diet: A ReviewDocument19 pagesHealth Implications of A Vegetarian Diet: A ReviewLindaNo ratings yet
- Probiotics and Health: A Review of The Evidence: E. WeichselbaumDocument35 pagesProbiotics and Health: A Review of The Evidence: E. WeichselbaumJavierjongNo ratings yet
- DR Sinatra HEART HEALTH NUTRITION PDFDocument8 pagesDR Sinatra HEART HEALTH NUTRITION PDFAnonymous BOXUxaNo ratings yet
- 10 Blood Tests For Vegans & VegetariansDocument3 pages10 Blood Tests For Vegans & VegetariansWeb WorldNo ratings yet
- Homocysteine and VitaminsDocument11 pagesHomocysteine and VitaminsHeart of the Valley, Pediatric CardiologyNo ratings yet
- Grades and Microbes: Ancient UnderstandingDocument3 pagesGrades and Microbes: Ancient UnderstandingKeesha Mae Urgelles TimogNo ratings yet
- The Silent Inflammation Zone Diet Reverses Chronic DiseaseDocument7 pagesThe Silent Inflammation Zone Diet Reverses Chronic DiseaseStanko VisnjicNo ratings yet
- A Critical Review of Coffee's Effects On Liver EnzymesDocument5 pagesA Critical Review of Coffee's Effects On Liver EnzymesMihai DragomirNo ratings yet
- mTOR - WikipediaDocument17 pagesmTOR - WikipediaNicholas FeatherstonNo ratings yet
- What is DetoxificationDocument6 pagesWhat is DetoxificationLaila AbdulNo ratings yet
- The Best Foods To Eat To Avoid Clogged Arteries The HealthyiuuihDocument2 pagesThe Best Foods To Eat To Avoid Clogged Arteries The Healthyiuuihrakebeef95No ratings yet
- The Brain Gut Nexus To DiseasesDocument44 pagesThe Brain Gut Nexus To DiseasesCarl MacCordNo ratings yet
- TheSecretofHydrogenRichWater Hidemitsu HayashiDocument3 pagesTheSecretofHydrogenRichWater Hidemitsu Hayashiiakhan45100% (1)
- Hydrazine CancerDocument12 pagesHydrazine CancerAnonymous yL790SIeHNo ratings yet
- The basics of how solar panels workDocument8 pagesThe basics of how solar panels workBishal LamichhaneNo ratings yet
- Homocysteine Supreme Doc RTDocument4 pagesHomocysteine Supreme Doc RTdianeculikNo ratings yet
- Expert Talk Transcripts from The Regenerate Yourself MasterclassDocument15 pagesExpert Talk Transcripts from The Regenerate Yourself MasterclassIrvan DavitNo ratings yet
- Breathe Your Limitations Away FullDocument88 pagesBreathe Your Limitations Away FullNikolina Zaljubljena JajacNo ratings yet
- Selective Serotonin Reuptake Inhibitors: Naoe, Marco Paulo RDocument38 pagesSelective Serotonin Reuptake Inhibitors: Naoe, Marco Paulo RMarco Paulo Reyes NaoeNo ratings yet
- Kirk Man Beginners Guide WebDocument40 pagesKirk Man Beginners Guide WebSophia Airall-ThomasNo ratings yet
- Gaba and Tryptophan Cravings Anxiety Trudy Scott 2022Document8 pagesGaba and Tryptophan Cravings Anxiety Trudy Scott 2022siesmannNo ratings yet
- VIOME+E-book Microbiom PDFDocument43 pagesVIOME+E-book Microbiom PDFNaša DjecaNo ratings yet
- Diabetes Type 2 BasicsDocument44 pagesDiabetes Type 2 BasicsMasunji EmanuelNo ratings yet
- ADIET Digital Image Processing Question BankDocument7 pagesADIET Digital Image Processing Question BankAdarshNo ratings yet
- Air Wellness QRSDocument2 pagesAir Wellness QRSapi-3743459No ratings yet
- Progibb LV Plus PGR - Low Voc FormulationDocument2 pagesProgibb LV Plus PGR - Low Voc FormulationDodik Novie PurwantoNo ratings yet
- Guidance Notes Blow Out PreventerDocument6 pagesGuidance Notes Blow Out PreventerasadqhseNo ratings yet
- Baseline Program Rev 3A Presentation 25 July 2020Document24 pagesBaseline Program Rev 3A Presentation 25 July 2020Shakti Sourava RautrayaNo ratings yet
- Apollo TyresDocument78 pagesApollo TyresADITYA33% (3)
- Casio AP-80R Service ManualDocument41 pagesCasio AP-80R Service ManualEngkiong Go100% (1)
- KAC-8102D/8152D KAC-9102D/9152D: Service ManualDocument18 pagesKAC-8102D/8152D KAC-9102D/9152D: Service ManualGamerAnddsNo ratings yet
- Product ListDocument4 pagesProduct ListyuvashreeNo ratings yet
- WL 318 PDFDocument199 pagesWL 318 PDFBeckty Ahmad100% (1)
- JUPITER 9000K H1PreliminaryDocument1 pageJUPITER 9000K H1PreliminaryMarian FlorescuNo ratings yet
- Nikola Tesla Was Murdered by Otto Skorzeny.Document12 pagesNikola Tesla Was Murdered by Otto Skorzeny.Jason Lamb50% (2)
- Reflective Essay 4Document1 pageReflective Essay 4Thirdy AngelesNo ratings yet
- QP (2016) 2Document1 pageQP (2016) 2pedro carrapicoNo ratings yet
- Thermal BurnsDocument50 pagesThermal BurnsPooya WindyNo ratings yet
- Chemistry of FormazanDocument36 pagesChemistry of FormazanEsteban ArayaNo ratings yet
- Uhf Leaky Feeder Rev CDocument4 pagesUhf Leaky Feeder Rev CLuis Isaac PadillaNo ratings yet
- Ultrasonic Weld Examination ProcedureDocument16 pagesUltrasonic Weld Examination ProcedureramalingamNo ratings yet
- The Temple of ChaosDocument43 pagesThe Temple of ChaosGauthier GohorryNo ratings yet
- Oral Nutrition Support NotesDocument28 pagesOral Nutrition Support Notesleemon.mary.alipao8695No ratings yet
- Lyceum of The Philippines University Cavite Potential of Peanut Hulls As An Alternative Material On Making Biodegradable PlasticDocument13 pagesLyceum of The Philippines University Cavite Potential of Peanut Hulls As An Alternative Material On Making Biodegradable PlasticJayr Mercado0% (1)
- Xii Neet Chemistry Mcqs PDFDocument30 pagesXii Neet Chemistry Mcqs PDFMarcus Rashford100% (3)
- Conjoint Analysis Basic PrincipleDocument16 pagesConjoint Analysis Basic PrinciplePAglu JohnNo ratings yet
- EP - EngineDocument4 pagesEP - EngineAkhmad HasimNo ratings yet
- 中美两国药典药品分析方法和方法验证Document72 pages中美两国药典药品分析方法和方法验证JasonNo ratings yet
- Proceedings of The 16 TH WLCDocument640 pagesProceedings of The 16 TH WLCSabrinaNo ratings yet
- Qualitative Research EssayDocument9 pagesQualitative Research EssayMichael FoleyNo ratings yet
- Effect of Some Algal Filtrates and Chemical Inducers On Root-Rot Incidence of Faba BeanDocument7 pagesEffect of Some Algal Filtrates and Chemical Inducers On Root-Rot Incidence of Faba BeanJuniper PublishersNo ratings yet
- Revolutionizing Energy Harvesting Harnessing Ambient Solar Energy For Enhanced Electric Power GenerationDocument14 pagesRevolutionizing Energy Harvesting Harnessing Ambient Solar Energy For Enhanced Electric Power GenerationKIU PUBLICATION AND EXTENSIONNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesFrom EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesRating: 3.5 out of 5 stars3.5/5 (56)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceFrom EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNo ratings yet
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- Gathering Moss: A Natural and Cultural History of MossesFrom EverandGathering Moss: A Natural and Cultural History of MossesRating: 4.5 out of 5 stars4.5/5 (347)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationFrom EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationRating: 4 out of 5 stars4/5 (1)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- The Magic of Reality: How We Know What's Really TrueFrom EverandThe Magic of Reality: How We Know What's Really TrueRating: 4.5 out of 5 stars4.5/5 (106)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)