Professional Documents
Culture Documents
Flagelo Español OK
Uploaded by
Sindy BravoCopyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Flagelo Español OK
Uploaded by
Sindy BravoCopyright:
Available Formats
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.
jpg
Ver imagen en tamao completo.
Quite el Marco
Imgenes resultantes
www.ugr.es/.../Microbiologia/images/08flag2.jpg
478 x 480 - 28 KB
La imagen puede estar a escala y/o sujeta a derechos de autor.
Abajo encontrar la imagen en su contexto original en la pgina: www.ugr.es/~eianez/Microbiologia/08flagelos.htm
Actualizado el mircoles, 15 febrero 2006
NDICE:
FLAGELOS
1.1
INTRODUCCIN: LA MOVILIDAD EN LAS BACTERIAS
1.2
ASPECTOS MORFOLGICOS
1.3
COMPOSICIN QUMICA Y ESTRUCTURA
1.3.1
FILAMENTO
1.3.2
CODO O GANCHO
1.3.3
CORPSCULO BASAL
1.4
ENSAMBLAJE DEL FLAGELO
1.5
MOVIMIENTO FLAGELAR
1.5.1
DESCRIPCION DEL MOVIMIENTO DE UNA BACTERIA PERITRICA
1.5.2
MECANISMO DEL MOVIMIENTO ALEATORIO (EN AUSENCIA DE ESTMULO)
1.5.3
1.5.2.1
EL MOTOR ES ROTATORIO Y GIRA EN AMBOS SENTIDOS
1.5.2.2
COMPONENTES DEL MOTOR Y DEL CONMUTADOR
1.5.2.3
ASPECTOS ENERGTICOS
LAS TAXIAS
1.5.3.1
DEFINICIONES
1.5.3.2
TIPOS DE TAXIAS
1.5.3.3
ESTUDIO DE LA QUIMIOTAXIA Y DE LA ADAPTACIN
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (1 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
1.5.3.3.1 ESTMULOS DEL SISTEMA DE RECEPCIN,
TRANSDUCCIN DE SEAL Y ADAPTACIN
1.5.3.3.2
EL CICLO DE EXCITACIN-ADAPTACIN
FLAGELOS PERIPLSMICOS
FIMBRIAS O PELOS (= PILI)
3.1
FIMBRIAS ADHESIVAS
3.2
PELOS SEXUALES DE BACTERIAS GRAM-NEGATIVAS
PROSTECAS
TALLOS O PEDNCULOS
ENLACES
DIAPOSITIVAS
1 FLAGELOS
1.1 INTRODUCCIN: LA MOVILIDAD EN LAS BACTERIAS
Los procariotas capaces de moverse lo hacen por alguno de estos sistemas:
Por flagelos provistos de un motor rotatorio
Por pelos de tipo IV, que dan lugar a dos tipos de desplazamiento:
Por sacudidas o contracciones en ciertas especies (de los gneros Neisseria y Pseudomonas)
Movilidad social por deslizamiento sobre superficies slidas en mixobacterias
Por secrecin de sustancias mucosas a travs de toberas (conjuntos de poros especiales) en la superficie
celular: lo que da origen a deslizamiento en ciertas cianobacterias y en mixobacterias, a modo de patinaje de la
clula sobre el moco depositado sobre la superficie.
Por un mecanismo de trinquete, responsable del deslizamiento en especies del grupo CytophagaFlavobacterium. En este caso existen un doble juego de protenas, uno de membrana citoplsmica y otro de
membrana externa. Ambos juegos estn engarzados entre s probablemente a nivel del peptidoglucano. Las
protenas de membrana citoplsmica se desplazan, y a su vez provocan movimiento en las de membrana
externa. Todo ello se asemeja a una correa sin fin o cadena de un tanque, que al moverse en un sentido sobre
una superficie, impulsan a la clula en el sentido opuesto.
El mejor estudiado de todos esos sistemas es de los flagelos. Los flagelos eubacterianos tpicos son largos
apndices filamentosos extracelulares, helicoidales, responsables del desplazamiento en medios lquidos de la mayor
parte de las bacterias mviles. Aunque empleamos la misma palabra para designar a los orgnulos locomotores de
procariotas y eucariotas, ambos son totalmente diferentes, tanto en estructura como en mecanismo de funcionamiento:
El filamento del flagelo bacteriano, que constituye su porcin externa ms visible, consta generalmente de un
solo tipo de protena, y en l no se realiza ningn trabajo quimiomecnico.
El mecanismo del flagelo bacteriano es rotatorio, con un motor reversible (funciona en los dos sentidos de giro).
La energa que propulsa a este motor no es ATP ni ninguna otra molcula con enlaces energticos, sino que
deriva directamente del gradiente de protones (fuerza protn-motriz).
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (2 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
1.2 ASPECTOS MORFOLGICOS
En preparaciones en fresco a microscopa ptica normal no se pueden detectar los flagelos individuales, debido a
su extrema delgadez (estn ligeramente por debajo del lmite resolutivo del microscopio). El microscopio de
contraste de fases logra visualizar los penachos densos de flagelos de ciertas bacterias.
En cambio, la microscopa ptica de alta intensidad en campo oscuro, as como la microscopa fluorescente s
logra discernir los flagelos.
Pueden estudiarse fcilmente mediante impregnacin argntica en preparaciones previamente fijadas por
procedimientos suaves que no destruyan la estructura (alcohol-ter) y con mordientes que la engruesan
artificialmente: cido tnico, sales de Al y Cr. (remitimos al alumno a la correspondiente prctica).
Los flagelos se observan como filamentos helicoidales largos y finos. La longitud es variable (no est
determinada de modo fijo): de 5 a 10 m, pero su anchura o dimetro es constante y uniforme para cada especie: en
Escherichia coli es de 20 nm. El carcter helicoidal del filamento es propio e intrnseco: para cada especie los
parmetros de la hlice (longitud de onda y amplitud) son fijos y caractersticos.
Existe una serie de especies Gram-negativas (Vibrio, Photobacterium, Bdellovibrio) que poseen flagelos muy
engrosados, debido a que estn envueltos en una vaina que presenta continuidad con la membrana externa.
El patrn de flagelacin (es decir, el nmero y localizacin de los flagelos) vara entre especies, y reviste inters en la
determinacin taxonmica:
un solo flagelo: bacterias monotricas. Normalmente la insercin del flagelo (en bacterias bacilares) es polar o
subpolar.
dos o ms flagelos formando un penacho, normalmente en un polo: lofotricas. (por ejemplo, en Spirillum
volutans hay ms de 80 flagelos en el penacho).
dos penachos de flagelos, uno en cada polo: bacterias anfitricas.
flagelos repartidos por toda la superficie: bacterias peritricas. (Por ejemplo, Escherichia coli posee 10 flagelos,
mientras que Proteus mirabilis posee cientos, dando aspecto muy denso a la disposicin de stos).
Ciertas bacterias presentan flagelos de insercin lateral, bien en penachos densos (Selenomonas), bien
diseminados (Pectinatus).
Existe una serie de especies Gram-negativas (Vibrio, Photobacterium, Bdellovibrio) que poseen flagelos muy
engrosados, debido a que estn envueltos en una vaina que presenta continuidad con la membrana externa
1.2 COMPOSICIN QUMICA Y ESTRUCTURA
El flagelo eubacteriano es una de las estructuras procariticas ms complejas, formada por ms de 20 tipos de
protenas estructurales. Ya a microscopa electrnica se pueden distinguir tres partes o subestructuras:
filamento helicoidal largo; est conectado a un corto segmento curvado denominado...
codo o gancho, que a su vez est unido al...
corpsculo basal. Este corpsculo basal est inmerso en las envueltas (membrana citoplsmica y pared), y
consta esencialmente de un cilindro que ensarta 1 o 2 parejas de anillos. En los ltimos aos se ha visto que,
relacionado con este corpsculo existen ms subestructuras. En conjunto, este corpsculo ampliado que ahora
conocemos tiene varias funciones:
anclar el flagelo a las envueltas
suministrar el mecanismo del movimiento (un motor rotatorio que puede girar en ambos sentidos)
albergar la maquinaria de exportacin de subunidades de protenas para su ensamblaje en la estructura
flagelar acabada
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (3 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
Estructura de un flagelo de una bacteria
Gram-negativa
1.3.1
FILAMENTO
El filamento es una estructura cilndrica fina, hueca y rgida, con aspecto helicoidal. Est constituido por el arrollamiento
de miles de subunidades idnticas de una protena llamada flagelina. La flagelina es una protena globular
relativamente elongada, con pesos moleculares variados, segn las especies (desde unos 15 kDa en algunos Bacillus
hasta unos 62 kDa en algunas enterobacterias). Las subunidades de flagelina se disponen formando una matriz
cilndrica, en la que se distinguen 11 hileras cuasi-axiales (casi verticales) de subunidades; las hileras cuasi-axiales se
denominan fibrillas. El cilindro est hueco (deja un canal en su interior de unos 3 nm). El filamento es notablemente
rgido, de modo que durante el movimiento activo slo se producen pequeas deformaciones, pero sin afectar a los
parmetros de la hlice.
En el extremo distal del filamento existe un capuchn formado por un pentmero de una protena distinta
(HAP2). Entre el filamento y el codo existen dos discos estrechos con dos tipos de protenas (HAP3 y HAP1). Son
estructuras adaptadoras que permiten la correcta interaccin entre filamento y codo.
El filamento tiene un papel pasivo, actuando de manera anloga a la hlice de un barco. El movimiento de
rotacin del motor (situado, como veremos, en el corpsculo basal) se comunica (a travs del codo) al filamento
helicoidal rgido, lo que permite el avance de la bacteria.
Tanto la flagelina nativa aislada como los filamentos intactos son buenos antgenos, constituyendo el
denominado antgeno H flagelar, especfico para cada especie e incluso para cada estirpe o raza.
1.3.2
CODO O GANCHO
Es una estructura curvada, acodada, de 55 nm de longitud, y 22 nm de dimetro, que conecta el filamento al
corpsculo basal. Consta de unas 120 unidades de una protena elongada, distinta de la flagelina (42 kDa), dispuestas
igualmente en una matriz cilndrica de 11 fibrillas. Parece ser que el codo acta a modo de juntura universal o flexible
entre el filamento y el corpsculo basal.
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (4 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
Entre el codo y el filamento existen dos discos de protenas accesorias del codo (HAP1 y HAP3). Cada uno de
los discos consiste en dos giros de hlice, e intervienen en el control del ensamblaje del flagelo. Son estructuras
adaptadoras que permiten la correcta interaccin entre filamento y codo.
1.3.3
CORPSCULO BASAL
Entendido el corpsculo basal en su sentido actual (ms amplio de lo que sabamos hace unos aos), es una notable
estructura que, inmersa en la membrana citoplsmica y en la pared celular, posee varias funciones:
ancla el flagelo a la clula
est relacionada con la funcin del motor rotatorio y del conmutador (cambio del sentido de giro)
alberga el aparato para la secrecin y correcto ensamblaje de la mayor parte del flagelo
En Gram-negativas, consta de dos pares de anillos coaxiales atravesados por un cilindro central hueco, junto
con otra serie de subestructuras anejas a todo lo anterior:
Los dos anillos exteriores se denominan L y P, y estn relacionados respectivamente con la membrana externa y
con el peptidoglucano. (A veces estos dos anillos estn conectados por una pared cilndrica que enmascara la
porcin del cilindro central).
Los dos anillos interiores se denominan S y M: el S est en el espacio periplsmico, inmediatamente por encima
de la membrana citoplsmica, y el M est inmerso plenamente en dicha membrana. Como ambos anillos estn
muy juntos, a veces se alude a ellos como anillo MS.
Ya hacia el lado citoplsmico de la membrana, por debajo del anillo M (y en contacto con l) existe un quinto
anillo, llamado anillo C, en el que existen tres protenas. Una de ellas, (FliG) parece ser el elemento central del
rotor, mientras que las tres (FliG, FliM y FliN) estn implicadas en la conmutacin del sentido del giro del
motor: permiten que el motor pueda girar en sentido de las agujas del reloj y en el sentido contrario al de las
agujas del reloj.
Rodeando al corpsculo basal, a modo de empalizada cilndrica, existen subunidades de dos protenas integrales
de membrana: MotA y MotB. Su papel parece ser doble:
Constituyen el estator del motor. Se sabe que MotB se proyecta hacia el espacio periplsmico, y
probablemente establece contactos con el peptidoglucano.
Por otro lado forman una especie de canal por donde pueden entrar los protones al citoplasma, lo que
constituye la base del mecanismo que surte de energa al motor.
En el hueco interior formado por los anillos MS y C se aloja una serie de protenas implicadas en la secrecin de
los monmeros de componentes del flagelo.
Con todo ello, la hiptesis que se maneja hoy es que, con el carburante de protones, el rotor gira respecto del
estator. Este giro se comunica al cilindro central, que a su vez debe de estar bien engarzado con al menos alguno de
los otros cuatro anillos. El giro se transmite al codo y, de ste al filamento. Como el filamento es una hlice rgida, al
girar hace avanzar a la bacteria en el medio acuoso.
En Gram-positivas la estructura del corpsculo basal es ms sencilla: cilindro central y una sola pareja de anillos
(el M y el S). Probablemente el anillo S est conectado con el peptidoglucano (sera anlogo en realidad al anillo P de
las Gram-negativas).
1.4 ENSAMBLAJE DEL FLAGELO
En la construccin del flagelo bacteriano intervienen unas 50 protenas, entre protenas estructurales que
formarn parte de la estructura definitiva (unas 20), y protenas accesorias que slo sirven durante este ensamblaje
(unas 30). Debido a esta complejidad, y a que, adems, diversas partes del flagelo deben de interaccionar con las
envueltas bacterianas (membrana, pared), no es de extraar que este ensamblaje est bien ajustado ni que la sntesis
de las diversas piezas est sometida a un estricto control gentico.
El orden de ensamblaje es en su mayor parte lineal y secuencial: tiene lugar desde las subestructuras ms
proximales hasta las ms distales.
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (5 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
Los componentes integrales de membrana (excluyendo a las protenas Mot), el doble anillo MS y el anillo C
emplean la habitual ruta Sec (procesamiento de pre-protenas con escisin del pptido lder).
Pero la mayor parte de las protenas cuyo destino est ms all de la membrana citoplsmica usan una ruta
especial, que es una variante del denominado sistema III de exportacin. Como dijimos, el aparato para esa
exportacin se alberga en el hueco de los anillos MS. Este aparato va exportando ordenadamente las
subunidades de las distintas subestructuras, las cuales viajan en fila india a travs del canal interior continuo
(que a su vez se va construyendo) del cilindro codo filamento. Las subunidades se van aadiendo en el
correspondiente extremo distal de la subestructura en formacin, y evitan su escape al medio merced a unas
protenas de capuchn en dichos extremos distales.
Por ejemplo, el filamento en construccin siempre dispone en su extremo distal de un capuchn del
pentmero de la protena HAP2. Dicho capuchn es plano hacia el exterior, pero posee cinco patas hacia
abajo, que forman una especie de cavidad relativamente amplia. Cuando por el hueco interior viaja una
nueva unidad de flagelina (an sin terminar de plegar), al llegar a la cavidad, hay un cambio de
conformacin en las patas de HAP2 que a su vez favorece el plegamiento adecuado de la flagelina y su
insercin en el lugar correcto. Cada vez que llega una subunidad de flagelina, el pentmero sufre el
cambio conformacional, y para acomodar la nueva subunidad se desplaza rotacionalmente, es decir, en el
ensamblaje, el pentmero de HAP2 est girando continuamente.
1.5 MOVIMIENTO FLAGELAR
En este apartado nos vamos a plantear dos objetivos de estudio principales:
descripcin del movimiento por flagelos (apartados 1.5.1)
mecanismo del movimiento en ausencia de estmulos (1.5.2)
cmo se ve modificado el mecanismo flagelar bsico ante la llegada de estmulos ambientales captados en la
superficies celular (1.5.3).
1.5.1
DESCRIPCION DEL MOVIMIENTO DE UNA BACTERIA PERITRICA
Vamos a estudiar en primer lugar cmo se produce el movimiento en bacterias peritricas como Escherichia coli o
Salmonella tiphymurium.
1. En ausencia de estmulos, en un medio ambiente uniforme, se puede observar un movimiento tridimensional
aleatorio, formado por periodos de unos pocos segundos de natacin en linea recta o ligeramente curvada
(carreras o corridas), interrumpidos por breves episodios (dcimas de segundo) de un movimiento angular catico
de la bacteria (viraje o cabeceo), tras de lo cual la clula entra en una nueva fase de natacin en lnea aunque con
una nueva direccin.
2. En un gradiente espacial de un estmulo ambiental, la bacteria responde modificando el anterior patrn. Se
alteran las probabilidades relativas de carreras y de virajes, de modo que la bacteria prolonga estadsticamente los
periodos de natacin hacia la direccin favorable (es decir, acercndose hacia un estmulo positivo y alejndose de
uno negativo), y disminuye la frecuencia de cabeceo. De esta manera se obtiene una locomocin aleatoria pero
estadsticamente sesgada, que propicia el avance neto en la direccin favorable. Este movimiento se denomina
taxia.
3. El comportamiento txico dura un tiempo limitado, que oscila de segundos a minutos, dependiendo de la
naturaleza e intensidad del gradiente. Tras la fase de excitacin inicial y de taxia, la bacteria se va adaptando al
estmulo, de modo que regresa finalmente al patrn aleatorio de movimiento, sin avance neto en ninguna direccin.
As pues, en las prximas pginas veremos la base de estos fenmenos:
existencia de un rotor flagelar de tipo rotacional, que es reversible.
Veremos que la bacteria tiene mecanismos para detectar un gradiente espacial de un estmulo, y
comprobaremos que este mecanismo acta como si la bacteria estuviera detectando de hecho un gradiente
temporal.
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (6 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
Conectado con el sistema de deteccin de estmulos, hay un sistema que hace que se vuelva al patrn aleatorio
ante la persistencia del estmulo (mecanismo de adaptacin).
1.5.2
MECANISMO DEL MOVIMIENTO ALEATORIO (EN AUSENCIA DE ESTMULO)
Vamos a resumir lo que veremos con ms detalle a continuacin:
El motor es rotatorio y tiene dos estados: giro en sentido antihorario, es decir contrario a las agujas del reloj
(CAR), y sentido horario, igual al de las agujas del reloj (AR)
Componentes y funcionamiento del motor y del conmutador y su relacin con la movilidad por carreras y con las
volteretas
Aspectos energticos del motor: el carburante del motor es la disipacin de protones a travs de cierta parte del
corpsculo basal.
1.5.2.1
EL MOTOR ES ROTATORIO Y GIRA EN AMBOS SENTIDOS
Desde hace varios aos, por experimentos en los que las bacterias quedaban unidas a cubreobjetos por anclaje
a ellos de los filamentos flagelares, se dedujo que el motor flagelar funciona como una mquina rotacional
reversible, o sea, tiene dos sentidos: el de las agujas del reloj (AR) y el contrario (CAR).
Veamos a continuacin las relaciones que se dan entre estos estados funcionales del motor y los fenmenos de
rotacin en lnea y virajes bruscos.
La natacin en lnea (carrera) se debe a la rotacin continua, durante un periodo de unos segundos, del motor,
en sentido CAR. En las bacterias peritricas, las fuerzas hidrodinmicas y mecnicas hacen que los filamentos de
los distintos flagelos se enrollen formando un haz o penacho paralelo al eje longitudinal de la clula. En este
penacho, cada flagelo gira independientemente. El giro de los diversos filamentos helicoidales del penacho,
empuja a la clula, originando la natacin en linea recta, con una velocidad de unos 25 m/seg.
Cuando el motor gira en sentido inverso, o sea AR, la nueva carga torsional que hace que el flagelo cambie de
conformacin. Mientras est cambiando esa conformacin, cada filamento tira independientemente de un lado de
la clula, lo que hace que esta pegue una voltereta que la reorienta de modo aleatorio.
1.5.2.2
COMPONENTES DEL MOTOR Y DEL CONMUTADOR
Como ya dijimos, parece que la empalizada de protenas integrales Mot A y Mot B forman parte del estator
del motor. Esta empalizada rodea al doble anillo MS, y probablemente sobre esta base fija gira el rotor (FliG) situado
bajo el doble anillo MS. Por debajo de ste, el anillo C, con sus tres tipos de protenas (FliG, M y N) acta como
conmutador del sentido de giro. El giro se transmite solidariamente al cilindro central, de l al codo, y de ah al
filamento helicoidal.
Si no existiera sistema de conmutacin del sentido de giro, el motor girara siempre en sentido antihorario o sea,
el sentido intrnseco (por defecto) de rotacin del motor flagelar es el CAR. Pero el motor recibe la influencia del
sistema conmutador del anillo C, que de vez en cuando invierte el sentido de giro (AR). Como veremos luego, a su vez
el sistema conmutador recibe informacin sensorial lo que se supondr que se alteraren las proporciones de tiempo en
que el motor gira en un sentido o en otro.
1.5.2.3
ASPECTOS ENERGTICOS
En condiciones normales con disponibilidad de nutrientes energticos en el medio ambiente de la bacteria, la
forma de energa que alimenta el motor flagelar es la fuerza protn motriz (f.p.m.), es decir, el potencial
electroqumico de protones (y en las bacterias marinas y alcalfilas, la fuerza motriz de los iones Na+).
La hiptesis actual sobre la manera en que se acopla el flujo de protones al motor con la rotacin dice lo
siguiente: cuando los protones cruzan la membrana citoplsmica, se unen a un determinado asprtico de la protena
MotB, lo cual le provoca un cambio conformacional. Este cambio conformacional del estator hace que el motor realice
un paso de rotacin (una fraccin de un giro). Luego, ese protn sale de la protena MotB y entra al citoplasma, con lo
que se regenera la configuracin original del estator. Para que el motor haga un giro completo, se necesitan unos 1000
protones. A su vez, el motor gira a la vertiginosa velocidad de 1000 revoluciones por minuto. De todas formas, la
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (7 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
velocidad de rotacin es variable, y depende de la intensidad del gradiente de protones.
De ese modo, la bacteria nada a altas velocidades relativas, de hasta 100 m/seg., (si un coche corriera a la
misma velocidad relativarompera la barrera del sonido!).
1.5.3
LAS TAXIAS
1.5.3.1
DEFINICIONES
En ausencia de un gradiente de estmulo, el movimiento de cada clula es a base de perodos de 2-4 segs de
natacin separados por virajes. El movimiento en un gradiente de estmulo se logra variando la frecuencia de virajes: si
la concentracin de un atrayente aumenta, o la de un repelente disminuye, las clulas no viran tan frecuentemente
como lo haran en un entorno uniforme. Por lo tanto, el resultado es que nadan en la direccin favorable ms tiempo, y
este sesgo respecto de la natacin aleatoria hace que exista un movimiento neto en relacin con el gradiente.
Este mecanismo no es una taxia en sentido estricto (se le puede llama clinocinesis, aunque este trmino apenas
se emplea); es un 50% menos efectivo que una taxia autntica, pero probablemente requiere mucha menos inversin
en maquinaria sensorial y motora.
1.5.3.2
TIPOS DE TAXIAS
Se distinguen principalmente aerotaxia, fototaxia y quimiotaxia.
1) aerotaxia: respuesta de migracin ante un gradiente de oxgeno molecular.
Bacterias anaerobias aerotaxia negativa.
Bacterias microaerfilas atradas a tensiones ptimas de O2 (menores que la atmosfrica).
Todas las bacterias aerobias y anaerobias facultativas aerotaxia positiva.
La aerotaxia positiva, al menos en enterobacterias, y en bacterias purpreas en crecimiento heterotrfico, tiene
un mecanismo diferente del mecanismo txico hacia otros atrayentes qumicos: no existe en realidad un receptor
especfico del oxgeno. La seal estimulatoria consiste en la utilizacin del oxgeno como aceptor final de electrones en
las cadenas transportadoras respiratorias. Esto provoca una alteracin de la f.p.m., que de alguna manera provoca un
cambio que aumenta la probabilidad de giro del rotor en sentido CAR.
2) Las fototaxias de las bacterias fotosintticas anxicas dependen de un mecanismo similar: detectan un gradiente de
intensidad de luz, no mediante un receptor especial, sino por medio del transporte de ee- fotosinttico, que a su vez
provoca un cambio en el gradiente electroqumico de H+. Este cambio afecta al motor flagelar.
3) Las quimiotaxias, especialmente las de Enterobacterias, son las taxias mejor estudiadas. Las abordaremos a
continuacin con cierto detalle.
1.5.3.3
ESTUDIODE LA QUIMIOTAXIA
Cuando se coloca a una bacteria en un gradiente de estmulo qumico se observa a lo largo del tiempo una
respuesta con 3 fases distintas:
fase de latencia (que dura unos 0.2 seg.), en la que el patrn de rotacin no se modifica.
rpida excitacin (medida aqu como la probabilidad de encontrar al rotor girando en sentido CAR).
fase de adaptacin lenta, hasta que se restablece el patrn inicial.
El comportamiento de quimiotaxia se pone en marcha cuando un estmulo qumico determinado (un
quimioefector) se une en la superficie bacteriana a un receptor especfico, lo que a su vez desencadena un proceso
de transduccin intracelular de esta seal, que finalmente llega al complejo conmutador del corpsculo basal
flagelar, lo que cambiar las proporciones iniciales de giro en sentido CAR y AR. Si el quimioefector produce una taxia
positiva, se denomina quimioatrayente, mientras que si desencadena una taxia negativa se denomina quimiorrepelente.
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (8 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
El sistema sensorial de la bacteria, ante un gradiente espacial de atrayente o repelente, origina una migracin
neta (acercamiento o alejamiento, respectivamente) haciendo que la duracin de una carrera en la direccin
favorable sea mayor, y para ello influye sobre el conmutador flagelar binario que determina el sentido de
rotacin.
La bacteria capta ese gradiente espacial, detectndolo de hecho como si fuera un gradiente temporal
autogenerado por ella misma mientras va movindose. Como veremos, la bacteria compara, mediante el
mismo receptor de membrana que capt el estmulo, la concentracin actual de la sustancia con la que tena en
los segundos anteriores.
Adems, la bacteria vuelve al patrn aleatorio de movimiento tras varios segundos o minutos del contacto inicial
con el gradiente: existe un mecanismo de adaptacin al estmulo que, como veremos, implica una modificacin
covalente de los receptores.
Estos son los aspectos que vamos a pasar a considerar a continuacin.
1.5.3.3.1
ELEMENTOS DE RECEPCIN, TRANSDUCCIN DE SEAL y DE ADAPTACIN
En enterobacterias se ha estudiado bien la quimiotaxis dependiente de un tipo de protenas sensoras de
estmulos qumicos denominadas MCP, acrnimo ingls de protenas quimiotcticas aceptoras de metilo, o
simplemente transductores. En Escherichia coli se han identificado cinco MCP, y cada una responde a un conjunto
determinado de quimiatrayentes y/o quimiorrepelentes. Por ejemplo, la MCP llamada Tar detecta los atrayentes
asprtico y maltosa y los repelentes cobalto y nquel.
Todas las MCP son protenas integrales de membrana citoplsmica, con un dominio periplsmico preparado
para unirse a los quimioefectores y un dominio citoplsmico dotado a su vez de dos zonas funcionales: una de
ellas es la regin transductora que genera la seal intracelular que va a llegar al motor flagelar, y la otra es la regin
metilable, con varios glutmicos que pueden aceptar radicales metilo (-CH3).
Los transductores MCP transmiten informacin al flagelo por medio de una cascada de transduccin
intracelular de la seal, en la que estn implicadas seis protenas: CheA, CheW, CheY, CheZ, CheR y CheB.
CheA: Es una protenquinasa que se autofosforila en una His concreta cuando el MPC se une a un
quimioefector. Es el regulador central del sistema quimiotctico, ya que en su forma fosforilada (CheA-P) fosforila
a CheY y a CheB.
CheW: es necesaria para el acoplamiento entre CheA y el dominio citoplsmico del MCP.
CheY: la protena CheY es fosforilada en un determinado asprtico por la CheA-P, y en su forma CheY-P
interacciona con el conmutador del anillo C del flagelo, lo cual incrementa la probabilidad de una inversin del
giro desde el CAR al AR, con lo que aumenta la probabilidad de virajes.
CheZ: contrarresta la seal de viraje al facilitar la desfosforilacin de CheY-P en CheY.
CheR: es una metil-transferasa que cataliza la transferencia de grupos metilo desde la S-adenosil-metionina
(SAM) hasta los glutmicos del dominio citoplsmico del MCP.
CheB: cuando esta protena es fosforilada por la CheA-P, se convierte en una metilasa (CheB-P) que retira los
metilos previamente unidos a los glutmicos del MCP.
Como veremos, la memoria celular para que la bacteria detecte gradientes de concentracin en funcin del
tiempo es el resultado de la metilacin de las MCP por la CheR, y de su desmetilacin por CheB-P.
1.5.3.3.2
EL CICLO DE EXCITACIN-ADAPTACIN
1. En un ambiente neutro (ausencia de estmulo), la MCP tiene un nivel basal de metilacin (una media de un
solo glutmico metilado en su dominio citoplsmico). En esta situacin, la CheA tiene un nivel basal de fosforilacin,
que sirve para fosforilar a CheY y a CheB. El motor flagelar presenta un patrn de giro de unos cuantos segundos
en sentido CAR ( natacin en lnea recta), y breves perodos de sentido AR ( virajes), que es la base del
movimiento aleatorio de la bacteria. Ahora veremos cmo este flujo basal de informacin hacia el flagelo y de
metilacin del MCP se altera con una sustancia atrayente.
2. Cuando aadimos el atrayente, ste se une al dominio periplsmico del MCP, lo que provoca un cambio
conformacional que se transmite hasta el dominio citoplsmico, de modo que se produce una modificacin de la
regin efectora de este dominio que transmite la seal hacia el motor flagelar. El tiempo que tarda en llegar al
motor explica el periodo de latencia. Durante esta fase el nivel de metilacin no se altera, pero el cambio
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (9 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
conformacional deja activados a los glutmicos metilables.
3. Transduccin intracelular de la seal: el cambio conformacional del dominio citoplsmico tiene el efecto de
inhibir la activacin de la CheA, de modo que desciende el nivel basal de su autofosforilacin, lo que se traduce
en que se fosforilan menos molculas de CheY y de CheB. Al haber menos CheY-P, llegan menos seales al
conmutador flagelar de que cambie a sentido igual al de las agujas del reloj. Por lo tanto, se prolongan los
perodos de natacin en lnea recta (y es ms probable que la bacteria se acerque al estmulo qumico).
4. Mientras tanto, ha estado actuando la metiltransferasa (CheR), que ha ido aadiendo metilos a los
glutmicos del dominio citoplsmico de la MCP. Si pasa un cierto tiempo y el dominio periplsmico sigue
ocupado por el quimioatrayente, se alcanza un gran nivel de metilacin en ese dominio citoplsmico. Esto hace que
el dominio efector citoplsmico vuelva a una configuracin nula, o sea, deja de emitirse seal excitadora. A ello
colabora igualmente que el nivel de metilesterasa (la protena CheB-P, es decir la forma fosforilada) sea bajo. Esta
es la explicacin molecular de por qu ocurre al cabo de unos segundos la adaptacin lenta al estmulo. Como la
metilacin es relativamente lenta, esto significa que si en un determinado momento la concentracin actual de
quimioefector (detectada por el grado de ocupacin del dominio periplsmico) es similar a la concentracin unos
segundos antes (manifestada por el nivel de metilacin del dominio citoplsmico), la bacteria sabe que lleva ya un
cierto tiempo acercndose al estmulo. Si la metilacin se ha estabilizado a un nivel alto, se produce un cambio
conformacional por el que el MCP deja de emitir seal (estado nulo), y la bacteria se ha adaptado al estmulo.
Como ejercicio, veamos qu pasara si ponemos un repelente: la unin del repelente al MCP
provoca un cambio conformacional que hace que el dominio citoplsmico interaccione ahora con el
complejo CheWCheA de modo que se aumenta la tasa de autofosforilacin de CheA. Ahora esta
CheA-P fosforila tanto a CheY como a CheB. La CheY-P difunde a la base del flagelo e interacciona
con el conmutador (anillo C), y "da la orden" de que se cambie ms a menudo a giro en sentido de las
agujas del reloj (AR), lo que hace que la bacteria vire caticamente ms a menudo. Con ello
aumentan sus probabilidades de que encuentre una direccin de natacin ms favorable (en sentido
opuesto al gradiente), en cuyo caso tendramos un patrn similar a cuando se une un atrayente.
Mientras tanto, la CheR ha encontrado ms difcil acceder a los glutmicos, y el mayor nivel de la
metilesterasa (CheB-P) hace que se eliminen metilos.
Esquema del funcionamiento del sistema de
transduccin de la seal de un
quimioatrayente (lee el texto para una
explicacin completa)
2 FLAGELOS PERIPLSMICOS
Son un tipo de flagelos que presenta exclusivamente el grupo de las Espiroquetas. Estas bacterias Gramnegativas son extremadamente finas y de forma helicoidal. Estn compuestas de:
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (10 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
cilindro protoplasmtico, formado por el protoplasto rodeado de la capa de peptidoglucano. El sculo de
murena de este PG tiene forma helicoidal, y es responsable de la tpica morfologa de estas bacterias;
membrana externa;
entre el cilindro protoplasmtico y la membrana externa se encuentran los peculiares flagelos, insertados
subpolarmente y enrollados alrededor del cilindro. Estos flagelos se denominan flagelos periplsmicos (=
endoflagelos = fibrillas axiales).
Estructura: Cada flagelo, en s mismo, es similar al tipo de flagelo clsico ya estudiado:
corpsculo basal, con dos pares de anillos (excepto en Leptospira, con slo un par);
codo
filamento, a base de subunidades de flagelina.
Ahora bien, lo caracterstico es la disposicin de los flagelos que salen de cerca de cada extremo, en relacin con el
cuerpo celular:
Esquema de los flagelos periplsmicos
de las espiroquetas
En la mayora de las especies, cada flagelo se extiende a lo largo del eje bacteriano, paralelo a la
superficie del cilindro protoplasmtico, hasta alcanzar los 2/3 de la longitud total; esto se traduce en que
los flagelos que salen de cada extremo se solapan en la zona central (la excepcin la tenemos en el gn.
Leptospira, en que no se solapan).
Las distintas fibrillas provenientes de cada extremo discurren paralelas y cercanas. El conjunto de estas
fibrillas se manifiesta como filamento axial, recubierto por membrana externa.
Obsrvese en el esquema el corte transversal de Cristispira, con sus numerosas fibrillas (=flagelos) axiales, que
conjuntamente determinan un filamento axial prominente (cresta), envuelto, por supuesto, en membrana externa.
Papel de los endoflagelos en la movilidad de las espiroquetas
Los flagelos de estas bacterias, paralelos al eje longitudinal de la clula, estn anclados al cilindro
protoplasmtico, que es rgido (mientras que la membrana externa es flexible). Al girar los flagelos de los dos
extremos en el mismo sentido, obligan a girar al cilindro rgido en un sentido, y a la membrana externa en sentidos
contrarios. El efecto es que el cilindro protoplasmtico gira en torno del filamento axial formado por los flagelos
periplsmicos. Esta es la base de los distintos tipos de movimientos que podemos observar en estas bacterias:
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (11 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
En medios lquidos se mueven
por avance muy rpido a modo de torniquete (el cuerpo bacteriano se comporta como un sacacorchos)
se pueden ver tambin contorsiones, latigazos, etc.
Sobre la superficie de medios slidos:
por rodamiento de la hlice. Esto se debe a que el filamento axial confiere movimiento de rotacin a la
membrana externa, lo que se traduce en que la bacteria pueda rodar. (Tome el alumno un muelle,
depostelo sobre una mesa y dle un empujn).
Flexiones. Se provocan ondas propagables del cilindro. La bacteria puede avanzar a modo de las larvas
de las mariposas gemetras
Todos estos tipos de movimiento son adaptaciones evolutivamente conseguidas que permiten el rpido avance en
medios de alta densidad (fangos espesos, mucosas de los animales, etc), medios donde los flagelos tpicos ya no
pueden servir adecuadamente. La evolucin ha hecho que las espiroquetas se adapten, por su peculiar estructura y la
de sus flagelos, a medios muy viscosos: de hecho, se mueven con mayor rapidez a 300-500 cP que a 10-50 cP.
Presentan, adems taxias positivas hacia esos medios viscosos.
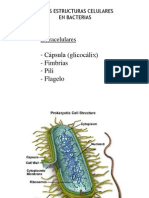
3 FIMBRIAS O PELOS (= PILI)
Son apndices filamentosos rectos y rgidos, ms cortos y ms finos (3-10 nm de dimetro) que los flagelos, y
que aparecen en muchas bacterias (sobre todo Gram-negativas). La mayora estn compuestos por un solo tipo
de protena (la pilina), de unos 17-25 kDa, cuyas subunidades se disponen en una matriz helicoidal que deja un
pequeo hueco central.
Estn implantados a nivel de membrana citoplsmica.
Nmero variable: desde 1 a varios cientos o miles por clula.
Disposicin: alrededor de todo el permetro celular, y a veces, de insercin polar.
Aislamiento: por simple agitacin mecnica de las clulas, seguido de ultracentrifugacin.
Composicin: ensamblaje de subunidades de pilina, protena globular muy hidrfoba.
Ensamblaje: insercin de subunidades de pilina en la base del pelo en crecimiento, a partir de pre-pilina, que
se procesa por escisin del correspondiente pptido-lder a su paso por la membrana citoplsmica.
Existen dos tipos principales de pili (pilus, en singular):
fimbrias adhesivas
pelos sexuales
3.1 FIMBRIAS ADHESIVAS
Son pelos de 4 a 7 nm de dimetro (segn especies), repartidas por toda la superficie y que funcionan como
adhesinas, es decir como estructuras para la adhesin a sustratos vivos o inertes. Condicionan varias propiedades
biolgicas, derivadas de sus propiedades adhesivas:
microcolonias y velos (los velos son pelculas formadas por acmulos de bacterias en medios estticos -en
reposo-).
adhesin a superficies inertes
adhesin superficies vivas. En el caso de bacterias patgenas, esta capacidad tiene que ver con su virulencia,
invasividad del tejido. Ejemplos de funcin como adhesinas:
en la formacin de la placa dental, por coagregacin de individuos de distintas especies sobre el diente;
factores de colonizacin de tejidos, por ejemplo en el gonococo (Neisseria gonorrhoeae) y en las cepas
uropatognicas de Escherichia coli.
La funcin de adhesina no reside en la pilina que constituye la inmensa mayora de la fimbria, sino en una protena
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (12 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
especial de la punta del pelo. La mayora de estas protenas de la punta pertenecen a la clase de las llamadas
lectinas, es decir, protenas o glucoprotenas capaces de unirse con gran afinidad a cadenas laterales de
polisacridos presentes en la membrana citoplsmica de las clulas del hospedador a las que se adhieren.
Aspectos genticos:
Los genes codificadores de las protenas de los pelos adhesivos son de localizacin cromosmica.
En muchas bacterias patgenas se da un fenmeno de variacin de fase: se trata de cambios rpidos y
reversibles por los que clulas piliadas (Fim+) pueden convertirse en no piliadas (Fim--), y viceversa. El
significado adaptativo es que los individuos Fim+ son los ms capaces de iniciar la colonizacin del animal
hospedador, pero son ms sensibles a la fagocitosis, mientras que una vez que han colonizado el epitelio, el
cambio a Fim-- permite que los individuos resistan mejor la fagocitosis.
Pero adems, existe otro interesante mecanismo adaptativo: las clulas Fim+ de algunas especies experimentan
fenmenos de variacin antignica, es decir, por una serie de mecanismos genticos, cambian la
especificidad antignica de su pilina, lo que supone una estrategia de evitacin contra el sistema inmune del
hospedador (un caso ms de esa carrera de armamentos evolutiva que se libra entre microorganismos y
organismos superiores).
Algunas pilinas (por ejemplo, en los gneros Moraxella y Neisseria) sirven tambin como parte del aparato
necesario para la transformacin gentica por ADN exgeno.
3.2 PELOS SEXUALES DE BACTERIAS GRAM-NEGATIVAS
Son ms largos y ms gruesos (unos 10 nm de dimetro) que las fimbrias adhesivas. Aparecen en menor
nmero (de 1 a 10 por clula), y su funcin es la de permitir los contactos iniciales en la conjugacin, como rgano
de reconocimiento entre la bacteria donadora, dotada del pelo sexual, y la receptora, carente de l. Sus genes son de
localizacin plasmdica.
Hay dos clases principales de pelos sexuales: los de de tipo F y los de tipo I, cada uno con un tipo de protena
distinta (genricamente conocida como pilina sexual). Son usados como receptores especficos por parte de algunos
fagos.
Hablaremos de los pelos sexuales cuando abordemos el captulo de Conjugacin bacteriana.
4 PROSTECAS
Son prolongaciones semirrgidas vivas, propias de ciertas bacterias, con un dimetro menor que el cuerpo
celular. Es decir, son apndices del cuerpo celular rodeados por membrana y pared celulares.
Funciones:
1) En algunas bacterias prostecadas funcionan en la reproduccin (prostecas reproductivas o hifas).
Ejemplos: Hyphomicrobium, Hyphomonas.
2) En Caulobacter acta como apndice para unin a sustratos inertes o para la formacin de rosetas de
varios individuos, merced al botn de anclaje situado en el extremo
3) En general, suponen un modo de aumentar la relacin superficie/volumen, lo cual redunda en:
a) mayor capacidad de flotabilidad para ciertas bacterias planctnicas;
b) mayor superficie para la captacin de nutrientes en ambientes oligotrficos.
Ejemplos: Ancalomicrobium, Prosthecomicrobium, Stella, etc.
5 TALLOS O PEDNCULOS
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (13 de 14)11-07-2008 10:14:04
Resultados Google para Imagen http://www.ugr.es/~eianez/Microbiologia/images/08flag2.jpg
Son estructuras filamentosas no vivas, terminadas en botones de anclaje (discos adhesivos), producidas por
secrecin continua de materiales polisacardicos en una zona concreta de la superficie bacteriana.
Funcin: Permite la unin de ciertas bacterias de hbitats acuticos a sustratos slidos, vivos o no.
Ejemplos:
Gallionella: es una bacteria con forma de media luna, de cuyo lado cncavo surgen de 3 a 40 filamentos
helicoidales, terminados en botones de anclaje.
Planctomyces: es una bacteria con forma de pera, con flagelo subpolar, que pasa parte de su vida anclada a
sustratos por medio de un tallo constituido por numerosas fibras rectas.
ENLACES
Para empezar, un artculo de una enciclopedia on-line que describe brevemente los flagelos procariotas y
eucariotas.
Interesantes vdeos sobre la movilidad por flagelos (Universidad de Harvard)
Para no perderse: El proyecto "nanomquina" nos asombra con unos espectaculares videos en 3-D sobre el
flagelo. El que trata sobre el ensamblaje es para quitarse el sombrero
El profesor Howard Berg escribe una estupenda pgina sobre el flagelo y la quimiotaxis, con figuras muy
instructivas
El profesor Macnab, uno de los mximos especialistas en flagelos bacterianos nos deleita con una revisin
centrada en el motor flagelar
Una serie de imgenes tridimensionales sobre el ensamblaje del flagelo
Una pgina sobre quimiotaxis mantenida por la Universidad de Bristol
Una pgina sobre el curioso movimiento por sacudidas, debido a pelos de tipo IV (con videos)
Una pgina sencilla sobre los distintos tipos de fimbrias y pelos (Universidad de Newcastle)
[ Introduccin e Historia ] [ Rasgos generales procariotas ] [ Tamao y forma ] [ Estructuras superficiales ]
[ Paredes procariotas ] [ Sntesis peptidoglucano ] [ Membrana y transporte ] [ Citoplasma y su contenido ]
[ Apndices filamentosos procariotas ] [ Diferenciaciones celulares ] [ Captacin de energa ] [ Nutricin microbiana ]
[ Ciclo celular y crecimiento ] [ Agentes fsicos ] [ Agentes qumicos ] [ Regulacin gnica ] [ Mutaciones en procariotas ]
[ Recombinacin. Transformacin ] [ Conjugacin ] [ Transduccin ]
Otras partes de esta web:
http://images.google.cl/imgres?imgurl=http://www....flagelo%2Bbacteriano%26gbv%3D2%26hl%3Des%26sa%3DG (14 de 14)11-07-2008 10:14:04
You might also like
- Informe Taller Resistencia MycobacteriumDocument18 pagesInforme Taller Resistencia MycobacteriumSindy BravoNo ratings yet
- Tecnicas AutoinmunesDocument27 pagesTecnicas AutoinmunesAgustin Garcia CanulNo ratings yet
- 1 Clase de Hemato, Hemostasia PrimariaDocument4 pages1 Clase de Hemato, Hemostasia PrimariaSindy BravoNo ratings yet
- Celulas T Reguladoras Infeccion y AutoinmunidadDocument8 pagesCelulas T Reguladoras Infeccion y AutoinmunidadSindy BravoNo ratings yet
- Informe 1 de InmunoDocument3 pagesInforme 1 de InmunoSindy BravoNo ratings yet
- Trabajo SociedadDocument2 pagesTrabajo SociedadSindy BravoNo ratings yet
- Entregable 3 Parcial 2 MicrobiologiaDocument3 pagesEntregable 3 Parcial 2 MicrobiologiaAlessia TéllezNo ratings yet
- Taller 4 MicrobiologiaDocument14 pagesTaller 4 MicrobiologiaAnder FrancoNo ratings yet
- Practica 1 - Quimiotaxis Bacteriana - PPTX (Reparado)Document16 pagesPractica 1 - Quimiotaxis Bacteriana - PPTX (Reparado)Angelica RojasNo ratings yet
- SensocentrismoDocument167 pagesSensocentrismoFlavio31No ratings yet
- Quimiotaxis bacteriana y eucariotaDocument6 pagesQuimiotaxis bacteriana y eucariotadavidclik09No ratings yet
- La QuimiotaxisDocument22 pagesLa QuimiotaxisMarisella Chumán Sánchez100% (3)
- RESUMEN Desarrollo y Morfología de NeutrófilosDocument7 pagesRESUMEN Desarrollo y Morfología de NeutrófilosErick2007No ratings yet
- Artículo Científico QuimiotaxisDocument7 pagesArtículo Científico QuimiotaxisTania Isabel Limongi GonzalezNo ratings yet
- Movimiento Celular FISIOLOGÍA (Guyton)Document11 pagesMovimiento Celular FISIOLOGÍA (Guyton)GeralKatiaCruzYufraNo ratings yet
- Clase 3-Microbiología AmbientalDocument36 pagesClase 3-Microbiología AmbientalJuan Carlos Malpartida SotoNo ratings yet
- Unidad 3. Caracterizacion MicrobianaDocument34 pagesUnidad 3. Caracterizacion Microbianadramireztimoteo_4192No ratings yet
- .Respuesta Inflamatoria Aguda. Consideraciones Bioquímicas y CelularesDocument16 pages.Respuesta Inflamatoria Aguda. Consideraciones Bioquímicas y CelularesFelipe Montiel MirandaNo ratings yet
- 3 Er Año Actividad Estimulo y RespuestaDocument9 pages3 Er Año Actividad Estimulo y Respuestasamanta barberisNo ratings yet
- 2 LeucocitosDocument67 pages2 LeucocitosRosmeri NavarroNo ratings yet
- 5 Flagelo y PiliDocument28 pages5 Flagelo y PiliAmari SolisNo ratings yet
- Factores de VirulenciaDocument1 pageFactores de VirulenciaJose Luis Reyes AragónNo ratings yet
- Ciclo CelularDocument10 pagesCiclo CelularLeninLeo 18No ratings yet
- 8° Guia de Clase No1-Biología-Funciones de Relacion-Segundo Periodo-2023Document10 pages8° Guia de Clase No1-Biología-Funciones de Relacion-Segundo Periodo-2023jaime londonoNo ratings yet
- Movimiento Microbiano y Respuestas SensorialesDocument12 pagesMovimiento Microbiano y Respuestas SensorialesIsabella MartinezNo ratings yet
- Clase 4 Capsula, Frimbia, Pili, FlageloDocument74 pagesClase 4 Capsula, Frimbia, Pili, Flageloapi-19878431100% (1)
- FLAGELODocument19 pagesFLAGELOConsuelo VargasNo ratings yet
- padA gen implicado en diferenciación DictyosteliumDocument136 pagespadA gen implicado en diferenciación DictyosteliumjhaebioNo ratings yet
- Informacion TaxiasDocument9 pagesInformacion TaxiasMiguel MANo ratings yet
- Todos Los CortesDocument197 pagesTodos Los CortesNini Sierra GutierrezNo ratings yet
- Fagocitosis: defensa celular contra agentes extrañosDocument20 pagesFagocitosis: defensa celular contra agentes extrañosJulio César Ramírez VenturaNo ratings yet
- Biometalurgia-Acidithiobacillus ThiooxidansDocument11 pagesBiometalurgia-Acidithiobacillus ThiooxidansJesús Alejandro Ochoa BerumenNo ratings yet
- Movilidad BacterianaDocument24 pagesMovilidad BacterianaCesar Eduardo Pastrano QuiñaNo ratings yet
- Capitulo 2 Estructura y Generalidades de BacteriasDocument15 pagesCapitulo 2 Estructura y Generalidades de BacteriasJosy ZambranoNo ratings yet
- Taller Dominio BacteriaDocument2 pagesTaller Dominio BacteriaKatia Racero100% (1)
- Las TaxiasDocument3 pagesLas Taxias0 0No ratings yet