You might also like
- 9 Desaturasa de Trypanosoma CruziDocument13 pages9 Desaturasa de Trypanosoma CruziAlexNo ratings yet
- Reporte 1Document7 pagesReporte 1GeorginaNo ratings yet
- α amilasaDocument10 pagesα amilasaDIANA BALDERASNo ratings yet
- Regulación Positiva de Leucina Aminopeptidasa-A en Raíces de Tomate Tratadas Con Cadmio - SpringerLinkDocument22 pagesRegulación Positiva de Leucina Aminopeptidasa-A en Raíces de Tomate Tratadas Con Cadmio - SpringerLinkDanteNo ratings yet
- Entamoeba HistolicaDocument4 pagesEntamoeba HistolicaJosepepe Bedolla BNo ratings yet
- Práctica 7 Actividad Enzimática de Las Peroxidasas.Document6 pagesPráctica 7 Actividad Enzimática de Las Peroxidasas.Agustin Pedraza GaleanaNo ratings yet
- Acción Enzimática InformeDocument25 pagesAcción Enzimática InformeNakiami ovNo ratings yet
- Bioquimica de Harvey 5ta EdicionDocument15 pagesBioquimica de Harvey 5ta EdicionmanuelNo ratings yet
- Actividad EnzimáticaDocument5 pagesActividad EnzimáticaTatiana LondoñoNo ratings yet
- Lipasas PapeleriaDocument5 pagesLipasas PapeleriaAntonio Marques GutierrezNo ratings yet
- Estrategia Catalítica de La LisozimaDocument17 pagesEstrategia Catalítica de La LisozimaLudwig Omar Julca Salazar100% (4)
- Fosfatasa ácida y su uso en el diagnóstico del cáncer de próstataDocument4 pagesFosfatasa ácida y su uso en el diagnóstico del cáncer de próstataAlejandro LeonNo ratings yet
- Digestión Enzimática Del Almidón Bioquimica FinalDocument7 pagesDigestión Enzimática Del Almidón Bioquimica FinalRoseline Galiano43% (7)
- Catalasa 10Document6 pagesCatalasa 10Emily BustillosNo ratings yet
- Actividad Enzimática de Peroxidasas VegetalesDocument8 pagesActividad Enzimática de Peroxidasas VegetalesEdgar Martin Hernandez HuaripaucarNo ratings yet
- Informe 5 Actividad Enzimatica FinalDocument7 pagesInforme 5 Actividad Enzimatica FinalAlirio BuelvasNo ratings yet
- Articulo Quimica OrganicaDocument10 pagesArticulo Quimica OrganicaLuis CutipaNo ratings yet
- Biomarcadores en anfípodos expuestos a sedimentosDocument13 pagesBiomarcadores en anfípodos expuestos a sedimentosroraysi_cm01No ratings yet
- 6 y 7 Inf. 1Document21 pages6 y 7 Inf. 1Yoselin Jazmin Azabache AbdhalaNo ratings yet
- RESUMENDocument5 pagesRESUMENIsabella JiménezNo ratings yet
- Info HidrolisisDocument4 pagesInfo HidrolisisJamie Vizcarra PalaciosNo ratings yet
- Clase Practica 12 Kenya Gil SDocument5 pagesClase Practica 12 Kenya Gil SKenya SantacruzNo ratings yet
- A04v32n4 PDFDocument7 pagesA04v32n4 PDFIsaac RivasNo ratings yet
- Lignina PeroxidasaDocument8 pagesLignina PeroxidasaViviana PérezNo ratings yet
- Limpio y Plegado Paper TraducidoDocument16 pagesLimpio y Plegado Paper TraducidoVicky W.No ratings yet
- Calderon-Flores LA U5A1Document10 pagesCalderon-Flores LA U5A1Lesly Adriana Calderon FloresNo ratings yet
- Inhibición No Competitiva de La Acetilcolinesterasa Por Alcaloides de Bromotirosina - Articulo Bioproceso EspañolDocument6 pagesInhibición No Competitiva de La Acetilcolinesterasa Por Alcaloides de Bromotirosina - Articulo Bioproceso EspañolFabian CelisNo ratings yet
- Universidad Popular de La Chontalp1Document8 pagesUniversidad Popular de La Chontalp1Cinthia Yarabeth Velázquez LópezNo ratings yet
- Informe BioquímicaDocument6 pagesInforme BioquímicaValeria AmpudiaNo ratings yet
- Actividad Antioxidante en Extractos de Algas MarinasDocument7 pagesActividad Antioxidante en Extractos de Algas MarinasCedricDiehlNo ratings yet
- Cinética (6) Cinética Enzimática de Hidrólisis Del Almidón Por Acción de La Diastasa-1Document9 pagesCinética (6) Cinética Enzimática de Hidrólisis Del Almidón Por Acción de La Diastasa-1Liz OtiosNo ratings yet
- Informe AmilasaDocument6 pagesInforme AmilasasandraNo ratings yet
- Mecanismos de acción del arsénicoDocument13 pagesMecanismos de acción del arsénicorosanna20147No ratings yet
- Actividad enzimática de la succinato deshidrogenasa y α-amilasa salivalDocument17 pagesActividad enzimática de la succinato deshidrogenasa y α-amilasa salivalLizeth VargasNo ratings yet
- Actividad enzimática succinato DHDocument7 pagesActividad enzimática succinato DHJair NavarreteNo ratings yet
- Fosfolipasa Taller BioquimicaDocument5 pagesFosfolipasa Taller BioquimicaAngélica María González SánchezNo ratings yet
- Preguntas Cortas Estructura BioquímicaDocument6 pagesPreguntas Cortas Estructura BioquímicafuggiaNo ratings yet
- INFORME4 GlicosidoscardiotonicosDocument8 pagesINFORME4 GlicosidoscardiotonicosAndrea AgudeloNo ratings yet
- Hongos de Podredumbre BlancaDocument20 pagesHongos de Podredumbre BlancaDiana Laritza VanegasNo ratings yet
- Efectos Atrazina Salud MedioambienteDocument31 pagesEfectos Atrazina Salud Medioambientekucolfmosteyrin100% (1)
- TP N°2 Enzimas 2022Document6 pagesTP N°2 Enzimas 2022Carina BrizzioNo ratings yet
- Determinación de la actividad de la α-amilasa salivalDocument10 pagesDeterminación de la actividad de la α-amilasa salivalCinthia Graciela Mejia EstradaNo ratings yet
- Articulo 3 - Un Nuevo Alcaloide de Quinolina de Las Raíces de Dictamnus Angustifolius - Seminario Te TesisDocument3 pagesArticulo 3 - Un Nuevo Alcaloide de Quinolina de Las Raíces de Dictamnus Angustifolius - Seminario Te TesisJuan AspilcuetaNo ratings yet
- Calb Enzima PDFDocument15 pagesCalb Enzima PDFJuan OlivaNo ratings yet
- Informe AmilasaDocument9 pagesInforme AmilasaJohanaS21No ratings yet
- Identificación BiomoléculasDocument12 pagesIdentificación BiomoléculasIsabella TrianaNo ratings yet
- Amilasa y LipasaDocument13 pagesAmilasa y LipasaKenia FragosoNo ratings yet
- Informe 4Document10 pagesInforme 4Claudia Lloclla G100% (1)
- Actividad Enzimatica de La Catalasa LaboratorioDocument15 pagesActividad Enzimatica de La Catalasa Laboratorioanderson0% (1)
- Enzimas digestivas amilasa y lipasaDocument9 pagesEnzimas digestivas amilasa y lipasaLiz AvalosNo ratings yet
- Clasificación de EnzimasDocument24 pagesClasificación de EnzimasSarha PuthNo ratings yet
- Actividad de Las Enzimas - Informe 4 - BiologiaDocument28 pagesActividad de Las Enzimas - Informe 4 - BiologiaSofia MerchanNo ratings yet
- Vocabulario de términos de biología molecular y bioquímicaDocument13 pagesVocabulario de términos de biología molecular y bioquímicaborrego2012No ratings yet
- Enzimologia ClinicaDocument18 pagesEnzimologia ClinicaRoberto Castellanos100% (1)
- Alcaloides Esteroidales de AsDocument5 pagesAlcaloides Esteroidales de AsMarco TorresNo ratings yet
- Caracterización Bioquímica de Extractos AcuososDocument18 pagesCaracterización Bioquímica de Extractos AcuososTalles FariaNo ratings yet
- Efecto Del PH y Temperatura Sobre La ActDocument10 pagesEfecto Del PH y Temperatura Sobre La ActRoberto MesíaNo ratings yet
- 25916-Article Text-40761-1-10-20190820Document15 pages25916-Article Text-40761-1-10-20190820ANDRES SARMIENTONo ratings yet
- ConstructivistaDocument10 pagesConstructivistaalfafriendNo ratings yet
- Magia de Las HierbasDocument34 pagesMagia de Las HierbasCocó Portugal89% (18)
- ConstantinoZoocriaMariposas PDFDocument25 pagesConstantinoZoocriaMariposas PDFJorge Antonio Peñaherrera CampoverdeNo ratings yet
- Ertmer, P. A., & Newby, T. J. 1993 (Traducción de Performance Improvement Quarterly Por Ferstadt & Szczurek) - Conductismo, Cognitivismo y Constructivismo...Document27 pagesErtmer, P. A., & Newby, T. J. 1993 (Traducción de Performance Improvement Quarterly Por Ferstadt & Szczurek) - Conductismo, Cognitivismo y Constructivismo...dani_elaa100% (2)
- Métodos de EsterilizaciónDocument6 pagesMétodos de EsterilizaciónJuan Villicaña DapénzxuNo ratings yet
- Higiene de Alimentos PDFDocument387 pagesHigiene de Alimentos PDFMundo Ingenieril75% (8)
- 1.portada de Cuaderno PDDocument1 page1.portada de Cuaderno PDJorge Antonio Peñaherrera CampoverdeNo ratings yet
- Micro Hongos Diversidad de Enzimas y Aplicaciones BiotecnologícasDocument6 pagesMicro Hongos Diversidad de Enzimas y Aplicaciones BiotecnologícasJorge Antonio Peñaherrera CampoverdeNo ratings yet
- Educacion Artistica Lineamientos Curriculares 1ro de BachilleratoDocument35 pagesEducacion Artistica Lineamientos Curriculares 1ro de BachilleratoJorge Antonio Peñaherrera CampoverdeNo ratings yet
- Azucares Re Duct Ores DNSDocument4 pagesAzucares Re Duct Ores DNSAraceli CastañedaNo ratings yet
- Botanica - Agricultura La Huerta Facil - Guia Practica Tomo I (C)Document33 pagesBotanica - Agricultura La Huerta Facil - Guia Practica Tomo I (C)debygnochi100% (22)
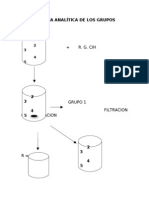
- Marcha Analítica de Los GruposDocument27 pagesMarcha Analítica de Los GruposJorge Antonio Peñaherrera CampoverdeNo ratings yet
- DanzaDocument14 pagesDanzaRonald Alexander Sayago MartinezNo ratings yet
- El arte del repujado en cobreDocument4 pagesEl arte del repujado en cobreJorge Antonio Peñaherrera CampoverdeNo ratings yet
- El Acrílico y Sus AplicacionesDocument1 pageEl Acrílico y Sus AplicacionesSebastian ZapataNo ratings yet
- 10 - Ejercicios Resueltos RedoxDocument11 pages10 - Ejercicios Resueltos RedoxMaximiliano Tlatlitlú83% (174)
- 06 Aves - Tipos de PicoDocument11 pages06 Aves - Tipos de PicoJorge Antonio Peñaherrera CampoverdeNo ratings yet
- Dominios de La VidaDocument3 pagesDominios de La VidaYenifer BeleñoNo ratings yet
- Rothia Mucilaginosa PDFDocument1 pageRothia Mucilaginosa PDFVictor PANo ratings yet
- Shigelosis: causas, síntomas e infección por ShigellaDocument134 pagesShigelosis: causas, síntomas e infección por ShigellaLorifloriNo ratings yet
- Lab. Microbiologia Lizeth Gabriela OspinaDocument10 pagesLab. Microbiologia Lizeth Gabriela OspinaLIZETH GABRIELA OSPINA SALAZARNo ratings yet
- Informe de Los BiofilmsDocument11 pagesInforme de Los BiofilmsAnghelo Tullume UriarteNo ratings yet
- El Origen de Las Células EucariontesDocument16 pagesEl Origen de Las Células EucariontesMacarena FranchiNo ratings yet
- Bacterias Gram Positivas y Gram NegativasDocument3 pagesBacterias Gram Positivas y Gram NegativasLa Toronja LokaNo ratings yet
- Origen de Los Seres VivosDocument12 pagesOrigen de Los Seres VivosNecrotemor NecrosNo ratings yet
- Primer Hemi MicroDocument17 pagesPrimer Hemi MicroMarjorie StefanieNo ratings yet
- Taxonomia Clasificacion EstructuraDocument38 pagesTaxonomia Clasificacion EstructuraJheremy Chacon RamirezNo ratings yet
- DIVERSIDAD CELULAR INFORME (Elodea)Document44 pagesDIVERSIDAD CELULAR INFORME (Elodea)Néstor R. Silva100% (1)
- EucariogénesisDocument17 pagesEucariogénesisManuelOyagueNo ratings yet
- Guia 6° Basico Celula y Teoria Celular. Con RespuestasDocument4 pagesGuia 6° Basico Celula y Teoria Celular. Con RespuestasEstefania SepulvedaNo ratings yet
- Guía de trabajos prácticos de microbiologíaDocument53 pagesGuía de trabajos prácticos de microbiologíaFernando Amblódegui GarcíaNo ratings yet
- 1 Sistema Digestivo-Circulatorio-Respiratorio-Excretor 2021-IiDocument12 pages1 Sistema Digestivo-Circulatorio-Respiratorio-Excretor 2021-IiZero MadsNo ratings yet
- Tendencias en Patógenos Bacterianos en Infecciones Del Tracto Respiratorio InferiorDocument27 pagesTendencias en Patógenos Bacterianos en Infecciones Del Tracto Respiratorio InferiorDiego Arriagada SantibáñezNo ratings yet
- Tema 11 - RELACIÓN HUÉSPED - BACTERIADocument15 pagesTema 11 - RELACIÓN HUÉSPED - BACTERIAGARCIA LITERATUSNo ratings yet
- Metodos de Preparación, Fijacion y Tecnicas de Coloración Simple, Tincion Diferencial y Selectivas.Document12 pagesMetodos de Preparación, Fijacion y Tecnicas de Coloración Simple, Tincion Diferencial y Selectivas.jorge briñez quesadaNo ratings yet
- Plata Coloidal Curar Sin AntibioticosDocument4 pagesPlata Coloidal Curar Sin AntibioticosMaríaNo ratings yet
- T033 47878779 TDocument128 pagesT033 47878779 Troshmel asenciosNo ratings yet
- Manual Torres Serv AuxDocument42 pagesManual Torres Serv AuxDavid BenitezNo ratings yet
- Sistema avanzado para recolección y transporte de muestras microbiológicasDocument8 pagesSistema avanzado para recolección y transporte de muestras microbiológicasneoegoNo ratings yet
- Vino y Salud Tesis Final PDFDocument30 pagesVino y Salud Tesis Final PDFPablo Montes StewartNo ratings yet
- El Reino Monera Cianobacterias para Segundo de SecundariaDocument5 pagesEl Reino Monera Cianobacterias para Segundo de SecundariaIvanAntonioVilchezNo ratings yet
- Gloria MontenegroDocument115 pagesGloria MontenegroLidia GomezNo ratings yet
- 5 - Ficha Técnica FOC R4T Actualización Abril 2021Document25 pages5 - Ficha Técnica FOC R4T Actualización Abril 2021yasserNo ratings yet
- Variedad de Medios de CultivoDocument2 pagesVariedad de Medios de CultivoDiego Ramirez100% (1)
- Los Seres Vivos. Taxonomía y Nomenclatura. Los Cinco Reinos, Relaciones Evolutivas. Los Virus y Su Patología. Otras Formas AcelularesDocument21 pagesLos Seres Vivos. Taxonomía y Nomenclatura. Los Cinco Reinos, Relaciones Evolutivas. Los Virus y Su Patología. Otras Formas Acelularesvictor gilNo ratings yet
- Membrana y estructuras bacterianas claveDocument45 pagesMembrana y estructuras bacterianas claveJorge Luis Ludeña PomaNo ratings yet
- Productos Mínimamente Procesados (IV Gama) DylanDocument25 pagesProductos Mínimamente Procesados (IV Gama) DylanHans Contreras WaissNo ratings yet