You might also like
- DigoxinaDocument7 pagesDigoxinaMari MoralesNo ratings yet
- GLUCÓLISISDocument2 pagesGLUCÓLISISdavo villaNo ratings yet
- Historia Natural de Diabetes InsipidaDocument1 pageHistoria Natural de Diabetes InsipidaKarim VargasNo ratings yet
- Metabolismo IntermedioDocument15 pagesMetabolismo IntermedioAdrian LeónNo ratings yet
- Valtek GlucosaDocument1 pageValtek GlucosaAlexandra Carbonel JulianNo ratings yet
- Oxigenoterapia de Alto Flujo PDFDocument5 pagesOxigenoterapia de Alto Flujo PDFCristian Manuel Goicochea ChuqueNo ratings yet
- Carbon ActivadoDocument5 pagesCarbon ActivadoDaniel Alejandro Rojas ToroNo ratings yet
- La Ruta de Las Pentosa-FosfatoDocument24 pagesLa Ruta de Las Pentosa-Fosfatoburgues1ta100% (1)
- RESPUESTA Metabolica Al Ayuno y Estres 2016Document45 pagesRESPUESTA Metabolica Al Ayuno y Estres 2016carlos garridoNo ratings yet
- Metabolismo de La Glucosa en Diferente OrganosDocument23 pagesMetabolismo de La Glucosa en Diferente OrganosStephany Fiorella Gómez RamírezNo ratings yet
- El Código ATC o Sistema de Clasificación AnatómicaDocument2 pagesEl Código ATC o Sistema de Clasificación AnatómicaMary Paez MorenoNo ratings yet
- AlopurinolMECACCIONDocument2 pagesAlopurinolMECACCIONMisael Ortega SNo ratings yet
- Práctica 3 PH y AmortiguadoresDocument8 pagesPráctica 3 PH y AmortiguadoresAldo MartínezNo ratings yet
- DESCRIPCION TeofilinaDocument17 pagesDESCRIPCION TeofilinaSuazo GloriaNo ratings yet
- Interacciones FarmacodinámicasDocument3 pagesInteracciones FarmacodinámicasJhordann Espinoza100% (1)
- Cetoacidosis Diabetica PDFDocument14 pagesCetoacidosis Diabetica PDFRoberto FajardoNo ratings yet
- Integración metabólica de carbohidratosDocument41 pagesIntegración metabólica de carbohidratospichicata0% (1)
- Estazolam insomnio tratamientoDocument10 pagesEstazolam insomnio tratamientoarmandoNo ratings yet
- Teicoplanina: mecanismo de acción, efectos y farmacocinética del antibiótico glicopéptidoDocument4 pagesTeicoplanina: mecanismo de acción, efectos y farmacocinética del antibiótico glicopéptidoAllison Velasquez MirandaNo ratings yet
- Universidad Autónoma de Chihuahua: Anova de Un FactorDocument15 pagesUniversidad Autónoma de Chihuahua: Anova de Un FactorKAREN HERNANDEZ MARTINEZNo ratings yet
- EPIRUBICINADocument4 pagesEPIRUBICINAmanuela AlejaNo ratings yet
- S300 EsDocument5 pagesS300 EsbinsjuniorNo ratings yet
- Etiologia de Litiasis Renal PDFDocument2 pagesEtiologia de Litiasis Renal PDFKelly0% (1)
- Bioquímica Notas Semana PesoDocument8 pagesBioquímica Notas Semana Pesoanelisse :3No ratings yet
- Libro BromatologiaDocument133 pagesLibro BromatologiaDelicia ColqueNo ratings yet
- Catabolismo carbohidratos glicolisis glucogenolisisDocument14 pagesCatabolismo carbohidratos glicolisis glucogenolisisSkuukzky baeNo ratings yet
- 06-3 Reacciones de Los CarbohidratosDocument40 pages06-3 Reacciones de Los CarbohidratosRicardo Alcántara ReyesNo ratings yet
- Regulacion de La Ruta de Las Pentosas FosfatoDocument9 pagesRegulacion de La Ruta de Las Pentosas FosfatoBlady SánchezNo ratings yet
- Suplementacion ConmicronitruentesDocument42 pagesSuplementacion ConmicronitruentesLuis Andrés Guaicha Alvarado100% (2)
- Neumonia 1Document6 pagesNeumonia 1cindy tellez amesquitaNo ratings yet
- Clasificación de Medicamentos.Document1 pageClasificación de Medicamentos.Fco ZárateNo ratings yet
- Programa PC 1Document37 pagesPrograma PC 1Carlos Fernando Soto AlonsoNo ratings yet
- Vancomicina Normon: ficha técnica de 500 mg y 1000 mg de vancomicinaDocument18 pagesVancomicina Normon: ficha técnica de 500 mg y 1000 mg de vancomicinaBeatriz PinedaNo ratings yet
- Tetracaina: anestésico local de acción prolongadaDocument3 pagesTetracaina: anestésico local de acción prolongadaNiinosk Páiz100% (1)
- CUESTIONARIODocument5 pagesCUESTIONARIOAbner Edgardo RodriguezNo ratings yet
- Farmacos SimpaticomimetricosDocument32 pagesFarmacos SimpaticomimetricosMadne cesi Gonzales melosichNo ratings yet
- Los EicosanoidesDocument14 pagesLos EicosanoidesalriccardiNo ratings yet
- Biosintesis y Degradacion de Nucleotidos 2009Document29 pagesBiosintesis y Degradacion de Nucleotidos 2009Leonardo Paricahua FloresNo ratings yet
- Acido PicricoDocument6 pagesAcido PicricodahanaNo ratings yet
- Cuestionario 7 CompletoDocument4 pagesCuestionario 7 CompletoDiego ANo ratings yet
- DifenhidraminaDocument6 pagesDifenhidraminaSam KrieguerichsNo ratings yet
- QUINIDINADocument13 pagesQUINIDINAAmairany ValdinNo ratings yet
- AligacionesDocument11 pagesAligacionesValentinMurilloTorresNo ratings yet
- Trimetoprim SulfometoxazolDocument4 pagesTrimetoprim SulfometoxazolMauricio Rueda RodriguezNo ratings yet
- Documento Completo de Cinetica EnzimaticaDocument11 pagesDocumento Completo de Cinetica EnzimaticaMonica VegaNo ratings yet
- AntihipertensivosDocument5 pagesAntihipertensivosCesar Alex Vera CariNo ratings yet
- Gestión de la calidad: Adiponectina, marcador de obesidad y resistencia a la insulinaDocument1 pageGestión de la calidad: Adiponectina, marcador de obesidad y resistencia a la insulinaromiraffin09No ratings yet
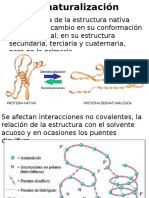
- DesnaturalizaciónDocument28 pagesDesnaturalizaciónErnesto GallegoNo ratings yet
- Prueba Del IndolDocument4 pagesPrueba Del IndolEddy Castellon RodriguezNo ratings yet
- Caso ABP Metabolismo de La Urea y Compuestos NitrogenadosDocument11 pagesCaso ABP Metabolismo de La Urea y Compuestos NitrogenadosNATALIA ROXANA CONCILCO JUAREZNo ratings yet
- Digestion AlmidonDocument12 pagesDigestion AlmidonRichardGildemeister100% (2)
- Digoxin ADocument15 pagesDigoxin AFLOR LOPEZNo ratings yet
- AbreviaturasDocument4 pagesAbreviaturasnashachrisNo ratings yet
- Enfoque Tradicional y Social de La SaludDocument14 pagesEnfoque Tradicional y Social de La Saludmaria alejandraNo ratings yet
- Péptidos: estructura, nomenclatura y funcionesDocument14 pagesPéptidos: estructura, nomenclatura y funcionesBelén Chamorro SevillaNo ratings yet
- Farmacos GastrointestinalesDocument38 pagesFarmacos GastrointestinalesErika MoyaNo ratings yet
- Antituberculosos PDFDocument18 pagesAntituberculosos PDFDaniel OrbeNo ratings yet
- LORAZEPAMDocument4 pagesLORAZEPAMLuis ValcarpNo ratings yet
- Bioquimica - 07 15Document36 pagesBioquimica - 07 15Lorena LisbethNo ratings yet
- Laboratorio #3. Unmsm Proteinas y AminoácidosDocument9 pagesLaboratorio #3. Unmsm Proteinas y AminoácidosMELISSA VERONICA TUDELANO OSPINANo ratings yet
- Plantilla de Funda Pikachu SentadoDocument1 pagePlantilla de Funda Pikachu SentadojhonatanNo ratings yet
- TítuloDocument6 pagesTítulojhonatanNo ratings yet
- Formato Apa GeneralDocument18 pagesFormato Apa GeneralJonathan' Cuotto Dellán'No ratings yet
- HormonasDocument1 pageHormonasjhonatanNo ratings yet
- HORMONASDocument1 pageHORMONASjhonatanNo ratings yet
- Danza y BaileDocument1 pageDanza y BaileJulio H H HipólitoNo ratings yet
- Danza y BaileDocument1 pageDanza y BaileJulio H H HipólitoNo ratings yet
- Anglicismos en El Mundo Del Deporte. Variación Lingüística y Sociolingüística (2012) PDFDocument25 pagesAnglicismos en El Mundo Del Deporte. Variación Lingüística y Sociolingüística (2012) PDFFernando PlansNo ratings yet
- La Toponimia Como Fuente para El Estudio Del Territorio (2009)Document9 pagesLa Toponimia Como Fuente para El Estudio Del Territorio (2009)Pablo FolgueiraNo ratings yet
- Alicia Misrahi - Los Poderes de VenusDocument375 pagesAlicia Misrahi - Los Poderes de VenusMirtha Euridice Rodriguez UrbinaNo ratings yet
- Conocer A DIOSDocument4 pagesConocer A DIOSOlaff Saavedra Ayala100% (1)
- Inicio Tesis MG USS 2015 (X)Document62 pagesInicio Tesis MG USS 2015 (X)Andrés Maturana FuentealbaNo ratings yet
- Cuadro Comparativo Karen Torres SocarrasDocument3 pagesCuadro Comparativo Karen Torres Socarraseverto gomezNo ratings yet
- Cadena EpidemiologicaDocument13 pagesCadena EpidemiologicaGian Ods100% (1)
- Eje 2 - Seminario de InvestigacionDocument10 pagesEje 2 - Seminario de InvestigacionLessly Zaray MurilloNo ratings yet
- Acta 1 29/05/2022Document3 pagesActa 1 29/05/2022Alejandro VillarroelNo ratings yet
- CULTURA CARAL, Chavin, ParacasDocument7 pagesCULTURA CARAL, Chavin, Paracasluis0316No ratings yet
- ELEMENTOS DE RETENCION Muros y Tablestac PDFDocument24 pagesELEMENTOS DE RETENCION Muros y Tablestac PDFWuilmerPeñaHuamanNo ratings yet
- Antecedentes de La InvestigacionDocument99 pagesAntecedentes de La InvestigacionJuan Yarleque VilcaNo ratings yet
- Jugando Con TíteresDocument2 pagesJugando Con TíteresDian Gianella Angela Rojas Quispe0% (1)
- Quiebre - Herramienta de Gestión para El Coach Ontológico Profesional (Daniel Rosales)Document7 pagesQuiebre - Herramienta de Gestión para El Coach Ontológico Profesional (Daniel Rosales)Laura RejoNo ratings yet
- Citas y Referencias Bibliograficas-IIIDocument6 pagesCitas y Referencias Bibliograficas-IIIMayumi Varas CernaNo ratings yet
- Ortografía 1. CorrecciónDocument2 pagesOrtografía 1. Correccióngespadas075No ratings yet
- La Ética de Nietzsche - Boulesis, Filosofía en La RedDocument4 pagesLa Ética de Nietzsche - Boulesis, Filosofía en La RedEric FigueroaNo ratings yet
- Ejercicios de La Pratica de Oscilaciones y MAS - FisicaDocument4 pagesEjercicios de La Pratica de Oscilaciones y MAS - FisicaGeraldNo ratings yet
- Secuencia de FuncionesDocument22 pagesSecuencia de FuncionesProf. Silvia MartinezNo ratings yet
- Marleny Rincon - Psicoterapia No 2Document2 pagesMarleny Rincon - Psicoterapia No 2Siau San VicenteNo ratings yet
- Agenda CristianaDocument221 pagesAgenda CristianaYanitza Angulo De Barreto100% (1)
- Contactos Mujeres Mujeres Maduras Buscan Sexo en MadridDocument2 pagesContactos Mujeres Mujeres Maduras Buscan Sexo en MadridConocer mujeres en MadridNo ratings yet
- 10 Errores en Tu Primer CapítuloDocument10 pages10 Errores en Tu Primer CapítuloArnul Aforismo100% (1)
- Complemento CircunstancialDocument30 pagesComplemento CircunstancialJaviera RodríguezNo ratings yet
- Análisis NuméricoDocument12 pagesAnálisis NuméricoOsvaldo AguirreNo ratings yet
- Investigación de La Desviación de Resultados MicrobiológicosDocument39 pagesInvestigación de La Desviación de Resultados MicrobiológicosSolutam MoraleNo ratings yet
- 2 BachilleratoDocument29 pages2 BachilleratoEzekiel HernándezNo ratings yet
- Profecia para Los Espiritus de NorteamericaDocument3 pagesProfecia para Los Espiritus de NorteamericaIván IllanesNo ratings yet
- Starobinski, Las Palabras Bajo Las Palabras, % Anagramas de SaussureDocument3 pagesStarobinski, Las Palabras Bajo Las Palabras, % Anagramas de SaussurerafelisarnNo ratings yet
- U1A1 - Ensayo Sobre La Importancia de La Negociacion. - TECNICASDocument21 pagesU1A1 - Ensayo Sobre La Importancia de La Negociacion. - TECNICASnanNo ratings yet