You might also like
- A Higher Level Classification of All Living OrganismsDocument14 pagesA Higher Level Classification of All Living OrganismsGaby Esquivel100% (1)
- Reguladores Proporcionales NFCDocument98 pagesReguladores Proporcionales NFCCarloMeztNo ratings yet
- Taxonomía - Wikipedia, La Enciclopedia LibreDocument713 pagesTaxonomía - Wikipedia, La Enciclopedia LibrejamilNo ratings yet
- Taxonomía - Wikipedia, La Enciclopedia LibreDocument33 pagesTaxonomía - Wikipedia, La Enciclopedia LibreLuciano SosaNo ratings yet
- Bienvenidos Al Curso de BiosistemáticaDocument47 pagesBienvenidos Al Curso de BiosistemáticaalexNo ratings yet
- TaxonomíaDocument33 pagesTaxonomíaGabriel Ch MendezNo ratings yet
- La TaxonomíaDocument13 pagesLa TaxonomíaOtto LópezNo ratings yet
- Niveles de Org y ClasificacionDocument8 pagesNiveles de Org y ClasificacionKentaroNo ratings yet
- TaxoDocument6 pagesTaxoAnonymous SJR6hq7No ratings yet
- Taxonomía AristitelesDocument66 pagesTaxonomía AristitelesDarly Bernabe Rimachi HuamanNo ratings yet
- 2°m Taxo 2Document7 pages2°m Taxo 2Jessica Lorena DíazNo ratings yet
- Definiciones TaxonomíaDocument30 pagesDefiniciones TaxonomíaFatima VargasNo ratings yet
- Clasificación de los seres vivos en 5 reinos y 3 dominiosDocument9 pagesClasificación de los seres vivos en 5 reinos y 3 dominiosAlexis GarciaNo ratings yet
- TaxonomíaDocument48 pagesTaxonomíaHelga DittusNo ratings yet
- Introducción a la taxonomía biológicaDocument12 pagesIntroducción a la taxonomía biológicaArgenis RamirezNo ratings yet
- Cuestionario de EcologiaDocument62 pagesCuestionario de Ecologiaolz2009No ratings yet
- TaxonomíaDocument36 pagesTaxonomíaniqmepaNo ratings yet
- TaxonomíaDocument21 pagesTaxonomíaCHAPZ RSELNo ratings yet
- InfotresDocument6 pagesInfotresPedro Abrahm Ramírez CondoriNo ratings yet
- Familia biologíaDocument9 pagesFamilia biologíakatherineNo ratings yet
- Texto-Clasificación de Los Seres VivosDocument6 pagesTexto-Clasificación de Los Seres VivosAdrianaNorteNo ratings yet
- Sistemas de clasificación taxonómicaDocument12 pagesSistemas de clasificación taxonómicaMiguel Rosas Martínez.No ratings yet
- GUÍA N°5 Biología 1° Medio Profesora Sussy SaavedraDocument6 pagesGUÍA N°5 Biología 1° Medio Profesora Sussy SaavedraNair ToscanoNo ratings yet
- TaxonomíaDocument21 pagesTaxonomíaedson choque bNo ratings yet
- 7 PDocument27 pages7 PJohan DionicioNo ratings yet
- TaxonomiaDocument15 pagesTaxonomiaalexandra leivaNo ratings yet
- Diferencia Entre Los Seres Vivos y Los Seres Inertes CCNN Sem 1Document6 pagesDiferencia Entre Los Seres Vivos y Los Seres Inertes CCNN Sem 1dave stanley iguasnia palominoNo ratings yet
- TAXONOMIA1234Document7 pagesTAXONOMIA1234prodiga de arcoNo ratings yet
- TaxonomiaDocument16 pagesTaxonomiaPABLONo ratings yet
- Patrones Biodiversidad J.C.E.Document9 pagesPatrones Biodiversidad J.C.E.Jenni ColinNo ratings yet
- Botanica Sistematica (Introducción)Document14 pagesBotanica Sistematica (Introducción)ItaloEvangelistaNo ratings yet
- Diapositiva - Importancia de La Taxonomia para El Conocimiento yDocument13 pagesDiapositiva - Importancia de La Taxonomia para El Conocimiento yVladimir Luciano AdamesNo ratings yet
- TAXONOMIADocument8 pagesTAXONOMIABraulio Calderon DediosNo ratings yet
- Texto 02Document9 pagesTexto 02NicanorNo ratings yet
- Revisions To The Classification, Nomenclature, An Diversity of Eukaryote. TranslatedDocument13 pagesRevisions To The Classification, Nomenclature, An Diversity of Eukaryote. TranslatedJazieel VázquezNo ratings yet
- La Clasificación de Los OrganismosDocument6 pagesLa Clasificación de Los OrganismosFisica Uno Conalep TolucaNo ratings yet
- TaxanomiaDocument8 pagesTaxanomiaAlexandra Junchaya NavarreteNo ratings yet
- TaxonomíaDocument14 pagesTaxonomíaSelene Ruiz LópezNo ratings yet
- TaxonomíaDocument29 pagesTaxonomíaLeoNo ratings yet
- Tema 1 TaxonomiaDocument6 pagesTema 1 TaxonomiaWilmer MendozaNo ratings yet
- Taxonomia BiologicaDocument12 pagesTaxonomia BiologicaDavid TujalNo ratings yet
- Caracteres TaxonómicosDocument11 pagesCaracteres TaxonómicosEkarine Gomez ReyezNo ratings yet
- Bioquimica 9Document8 pagesBioquimica 9carlos felipe bejaranoNo ratings yet
- TaxónDocument6 pagesTaxónAntonio TreminiNo ratings yet
- Guia Aprendizaje Estudiante 6to Grado Ciencia f3 s9Document7 pagesGuia Aprendizaje Estudiante 6to Grado Ciencia f3 s9Juan Carlos CórdovaNo ratings yet
- Sistema de Clasificación TaxonomicaDocument5 pagesSistema de Clasificación TaxonomicaSilvana SanchezNo ratings yet
- Grupos TaxonómicosDocument5 pagesGrupos TaxonómicosAngiee RoödzNo ratings yet
- GUÍA 1 TAXONOMÍA CompletaDocument15 pagesGUÍA 1 TAXONOMÍA CompletaMariana OspinaNo ratings yet
- Decimo Clase N 1Document9 pagesDecimo Clase N 1Homero AlvarezNo ratings yet
- Introduccinalataxonoma 20230827092936Document2 pagesIntroduccinalataxonoma 20230827092936daniela sanchezNo ratings yet
- Taxonomía de BacteriasDocument49 pagesTaxonomía de Bacteriasjuan perez100% (2)
- Guía sobre taxonomía y clasificación de organismosDocument32 pagesGuía sobre taxonomía y clasificación de organismosGustavo RangelNo ratings yet
- La TaxonomíaDocument8 pagesLa TaxonomíaElga AnaNo ratings yet
- La TaxonomíaDocument4 pagesLa TaxonomíaAlfredo MorenoNo ratings yet
- Lectura 1 Taxonomía y Biodiversidad PDFDocument15 pagesLectura 1 Taxonomía y Biodiversidad PDFyemeneses126No ratings yet
- Leandro TrosssaarDocument3 pagesLeandro Trosssaaresteban herreraNo ratings yet
- Clasificacion de Los Seres Vivos - Sistematica y Filogenia - ApunteDocument8 pagesClasificacion de Los Seres Vivos - Sistematica y Filogenia - ApunteAbi Lopez de AntuenoNo ratings yet
- PDF, Ciencias Naturales Grado SextoDocument12 pagesPDF, Ciencias Naturales Grado SextoAnggie Nathalie Tobar ContrerasNo ratings yet
- La TaxonomíaDocument2 pagesLa Taxonomíacamila velandiaNo ratings yet
- Catálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónFrom EverandCatálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónNo ratings yet
- Catálogo PISCINA 2023Document35 pagesCatálogo PISCINA 2023juanNo ratings yet
- Mecanismos de Transmisión Del MovimientoDocument3 pagesMecanismos de Transmisión Del MovimientoJESUS EMMANUEL RAYON FLORESNo ratings yet
- Parcial 2 - Andrea Michelle Tovar Fierro - 157277 PDFDocument4 pagesParcial 2 - Andrea Michelle Tovar Fierro - 157277 PDFMi Si HoNo ratings yet
- Proyecto de Maquina PropulsoraDocument97 pagesProyecto de Maquina PropulsoradannyNo ratings yet
- 100 Años, Emiliano LujánDocument12 pages100 Años, Emiliano LujánGobierno Autónomo Municipal de La PazNo ratings yet
- Nomenclatura de FenotiposDocument10 pagesNomenclatura de FenotiposEvelyn MoralesNo ratings yet
- wiki/: Motores de Combustión Interna Ciclo Otto Carburador Inyección de CombustibleDocument50 pageswiki/: Motores de Combustión Interna Ciclo Otto Carburador Inyección de CombustibleAlexander EstivenNo ratings yet
- Examen 03 de Aritmética para Segundo de PrimariaDocument8 pagesExamen 03 de Aritmética para Segundo de PrimariaSujel RojasNo ratings yet
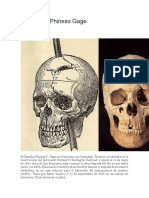
- El Caso de Phineas GageDocument8 pagesEl Caso de Phineas GageHevechito Palomino PadeNo ratings yet
- Análisis Textos PeriodísticosDocument1 pageAnálisis Textos PeriodísticosRosario Garcia MarinNo ratings yet
- Zonas Sismicas de MexicoDocument2 pagesZonas Sismicas de MexicoAlessander CabreraNo ratings yet
- 06 - Historia de La Creación (Original)Document19 pages06 - Historia de La Creación (Original)roberto ibanez100% (1)
- CuidaMedioAmbienteDocument2 pagesCuidaMedioAmbienteAlonso VigoNo ratings yet
- SAI Visiones Internas e InspiracionesDocument94 pagesSAI Visiones Internas e InspiracionesoronuevoNo ratings yet
- Invasion y Mestastasis 12Document17 pagesInvasion y Mestastasis 12Daniel Apaza ZegarraNo ratings yet
- PONS de Emergencias.Document8 pagesPONS de Emergencias.Antonio NietoNo ratings yet
- Bc-4763 Quiroz AlcantaraDocument252 pagesBc-4763 Quiroz Alcantarailder rosasNo ratings yet
- Tecnicas de Conteo Ejercicios ResueltosDocument18 pagesTecnicas de Conteo Ejercicios ResueltosavileitorNo ratings yet
- PRÁCTICA N1 Elt 2641Document17 pagesPRÁCTICA N1 Elt 2641JHONNY VILCAEZ CHAVARRÍANo ratings yet
- Glosario PetroDocument4 pagesGlosario PetroRufo WuarnerNo ratings yet
- Las Aplicaciones de La Presión OsmóticaDocument8 pagesLas Aplicaciones de La Presión OsmóticaJuleysy ValenzuelaNo ratings yet
- Taller 2 Reacciones CineticasDocument4 pagesTaller 2 Reacciones CineticasDiana S. VásquezNo ratings yet
- Ejercicios - García GarciaDocument9 pagesEjercicios - García GarciaRonald LlerenaNo ratings yet
- Actividad 3Document2 pagesActividad 3Arturo DreadNo ratings yet
- Etapas de Producción de Una EmpresaDocument3 pagesEtapas de Producción de Una EmpresaGuillermo SamuelNo ratings yet
- Cuestionario #12Document5 pagesCuestionario #12george guerraNo ratings yet
- Coeducación y perspectiva de géneroDocument2 pagesCoeducación y perspectiva de génerodianaNo ratings yet
- El InconsienteDocument8 pagesEl InconsienteABIGAIL HAZEL BAUTISTA TREJONo ratings yet
- Chacras Amazónicas - Guía para El Manejo de PlagasDocument156 pagesChacras Amazónicas - Guía para El Manejo de PlagasCarreño Rangel Tania YamileNo ratings yet
- Reporte Tipos de SueloDocument2 pagesReporte Tipos de SueloAbisail Peralta100% (3)