You might also like
- Diretrizes de laparoscopia urológicaFrom EverandDiretrizes de laparoscopia urológicaOscar Eduardo Hidetoshi FugitaNo ratings yet
- Aula 1 GeneralidadesDocument21 pagesAula 1 GeneralidadesjussaraNo ratings yet
- Resumo Neurofisiologia Clinica Principios Basicos e Aplicacoes Luiz Carlos PintoDocument2 pagesResumo Neurofisiologia Clinica Principios Basicos e Aplicacoes Luiz Carlos PintoJosehp Millares TolaNo ratings yet
- Telencéfalo AnatomiaDocument62 pagesTelencéfalo AnatomiaReinaldo RiquetNo ratings yet
- Diagnóstico diferencial para quadro de mal epilépticoDocument7 pagesDiagnóstico diferencial para quadro de mal epilépticoAndréaNo ratings yet
- Atlas Fotografico Neuro AnatomiaDocument27 pagesAtlas Fotografico Neuro AnatomiaRavenna Horana Alves da Silva100% (1)
- Caso - Doença de McArdleDocument14 pagesCaso - Doença de McArdleabresserNo ratings yet
- Roteiro Mód. XX - 2017-2 - PBL7Document13 pagesRoteiro Mód. XX - 2017-2 - PBL7levy_rebouçasNo ratings yet
- Mielomeningocele Atuação To 187 PDFDocument237 pagesMielomeningocele Atuação To 187 PDFMariana JoãoNo ratings yet
- Laboratório de MorfofuncionalDocument8 pagesLaboratório de MorfofuncionalpauloNo ratings yet
- Miastenia GravisDocument18 pagesMiastenia Gravisanasuhet100% (2)
- Hipoxia PerinatalDocument13 pagesHipoxia PerinataljulialeoNo ratings yet
- Abordagem Da Vertigem: "Tontura"Document2 pagesAbordagem Da Vertigem: "Tontura"Joana CoelhoNo ratings yet
- Questões de Neuroanatomofisiologia 1-20Document5 pagesQuestões de Neuroanatomofisiologia 1-20Kim MansurNo ratings yet
- Celulas Acidofilas-Parte 1Document4 pagesCelulas Acidofilas-Parte 1Diego MaciasNo ratings yet
- Questionário da Avaliação N1Document5 pagesQuestionário da Avaliação N1derciaNo ratings yet
- Atlas de NeuroimagemDocument44 pagesAtlas de NeuroimagemMarcelle BelgaNo ratings yet
- Funções Formação ReticularDocument22 pagesFunções Formação ReticularDra. MarciaNo ratings yet
- Anatomia Na Prática PDFDocument228 pagesAnatomia Na Prática PDFLarissa ReisNo ratings yet
- Roteiro Neuroanatomia Práticas 1 A 5 - 2023Document11 pagesRoteiro Neuroanatomia Práticas 1 A 5 - 2023Ellen Santos Rodrigues100% (1)
- Litiase UrinariaDocument8 pagesLitiase UrinariaLya_ANo ratings yet
- Anatomia do olhoDocument30 pagesAnatomia do olhoJulia RavaziNo ratings yet
- Aula 4 Sistema CardiovascularDocument23 pagesAula 4 Sistema CardiovascularNatália Chaves GondimNo ratings yet
- Shapiro 2Document12 pagesShapiro 2Yasmin CapriniNo ratings yet
- AULA 12 - Trauma GenitourinárioDocument79 pagesAULA 12 - Trauma GenitourinárioAvhaPaixãoNo ratings yet
- Neuroanato - Vascularização Do SNC e Barreiras EncefálicasDocument4 pagesNeuroanato - Vascularização Do SNC e Barreiras Encefálicasviniciusdays100% (1)
- Embrio - Embriologia Básica - Moore & Persaud (7ª Ed.)Document455 pagesEmbrio - Embriologia Básica - Moore & Persaud (7ª Ed.)santos.rdutraNo ratings yet
- 10 - Inervação Do PéDocument9 pages10 - Inervação Do PéRodrigo AmaralNo ratings yet
- Anatomia pulmonar e doenças do tabagismoDocument204 pagesAnatomia pulmonar e doenças do tabagismocjkcjk100% (1)
- Estudos Dirigidos de NeuroanatomiaDocument5 pagesEstudos Dirigidos de NeuroanatomiaLeonardo FinelliNo ratings yet
- Seio CavernosoDocument29 pagesSeio CavernosoTúlio Maranhão100% (1)
- Fisioterapia Neurológica GuiaDocument31 pagesFisioterapia Neurológica GuiaIdânia LopesNo ratings yet
- Sistema NervosoDocument94 pagesSistema NervosoIsabela LaynesNo ratings yet
- Encefalites Autoimunes - Abordagem Prática para SituaçõesDocument10 pagesEncefalites Autoimunes - Abordagem Prática para Situaçõesfernandosousa7No ratings yet
- O Sistema Nervoso Autônomo e a Medula AdrenalDocument3 pagesO Sistema Nervoso Autônomo e a Medula AdrenalJomar BorgesNo ratings yet
- Perfil e prevalência de tumores do SNCDocument6 pagesPerfil e prevalência de tumores do SNCAllan MaiaNo ratings yet
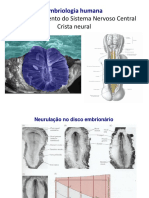
- Desenvolvimento do Sistema Nervoso CentralDocument30 pagesDesenvolvimento do Sistema Nervoso CentralBea1999100% (2)
- Material 3Document29 pagesMaterial 3Reinaldo RiquetNo ratings yet
- Anatomia do Sistema Esquelético Axial e TóraxDocument6 pagesAnatomia do Sistema Esquelético Axial e TóraxGabriela GonçalvesNo ratings yet
- Função do Sistema NervosoDocument8 pagesFunção do Sistema NervosoCristiana Leal Sabel100% (1)
- Histologia ClínicaDocument205 pagesHistologia ClínicaAdeilson ReisNo ratings yet
- Exercício Sobre Sistema NervosoDocument11 pagesExercício Sobre Sistema Nervosofernandam820No ratings yet
- Aula 3 - Córtex CerebralDocument18 pagesAula 3 - Córtex CerebralPamela SouzaNo ratings yet
- Vascularizaao Do Sistema Nervoso Central - CompressDocument70 pagesVascularizaao Do Sistema Nervoso Central - CompressVinicius Herbet100% (1)
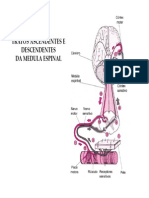
- Tratos Ascendentes e Descendentes Da ME (Modo de Compatibilidade)Document13 pagesTratos Ascendentes e Descendentes Da ME (Modo de Compatibilidade)Natália PimentelNo ratings yet
- Estruturas do sistema respiratórioDocument4 pagesEstruturas do sistema respiratórioSueniaNo ratings yet
- NEUROANATOMIA 10 - Macroscopia Do Diencéfalo - MED RESUMOS 2012 PDFDocument3 pagesNEUROANATOMIA 10 - Macroscopia Do Diencéfalo - MED RESUMOS 2012 PDFdellionkingNo ratings yet
- Funções Motoras Da Medula Espinhal e Do Tronco CerebralDocument15 pagesFunções Motoras Da Medula Espinhal e Do Tronco CerebralLeni BarrosNo ratings yet
- Fisiologia Aplicada e Psicobiologia 1Document8 pagesFisiologia Aplicada e Psicobiologia 1Camila BuenoNo ratings yet
- Anatomia LinfaticoDocument6 pagesAnatomia Linfaticorentcs100% (1)
- Trabalho NeuronioDocument10 pagesTrabalho NeuronioLETICIA MACEDO MAGGIORININo ratings yet
- Glossário AnatomiaDocument7 pagesGlossário AnatomiaCintia Flôr100% (2)
- LombalgiaDocument79 pagesLombalgiaSamuel BrancoNo ratings yet
- Irrigação ResumidaDocument3 pagesIrrigação Resumidaisabella cristinna da silva costaNo ratings yet
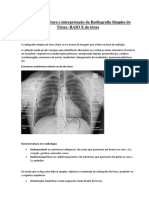
- Bases para Leitura e Interpretação Da Radiografia Simples Do TóraxDocument6 pagesBases para Leitura e Interpretação Da Radiografia Simples Do TóraxErnesto dos SantosNo ratings yet
- 208 686 1 PBDocument10 pages208 686 1 PBAna AraújoNo ratings yet
- 17021-Texto Do Artigo-72245-1-10-20100329Document6 pages17021-Texto Do Artigo-72245-1-10-20100329Ian JhemesNo ratings yet
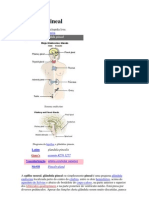
- Glândula PinealDocument11 pagesGlândula PinealJuliana July CripaNo ratings yet
- Atividade Diencefalo - FinalDocument3 pagesAtividade Diencefalo - FinalAndrea RebeloNo ratings yet
- Neurofisiologia Do Sono PDFDocument11 pagesNeurofisiologia Do Sono PDFYasmin Oliveira100% (1)
- LAPAROTOMIADocument28 pagesLAPAROTOMIAphoenixbeta100% (1)
- Habilidades para comunicação com crianças e adolescentesDocument3 pagesHabilidades para comunicação com crianças e adolescentesphoenixbetaNo ratings yet
- Refluxo Gastro EsofagicoDocument18 pagesRefluxo Gastro EsofagicoCaroline PizzaniNo ratings yet
- Bacteriuria AssintomaticaDocument10 pagesBacteriuria AssintomaticaphoenixbetaNo ratings yet
- Refluxo Gastro EsofagicoDocument18 pagesRefluxo Gastro EsofagicoCaroline PizzaniNo ratings yet
- Sem An ADocument1 pageSem An AphoenixbetaNo ratings yet
- MilleDocument2 pagesMillephoenixbetaNo ratings yet
- TutsDocument1 pageTutsphoenixbetaNo ratings yet
- CravingDocument7 pagesCravingphoenixbetaNo ratings yet
- Objetivos 04Document1 pageObjetivos 04phoenixbetaNo ratings yet
- CravingDocument7 pagesCravingphoenixbetaNo ratings yet
- Laudo!Visualizar DoDocument1 pageLaudo!Visualizar DoSara EsquedaNo ratings yet
- Tecido SanguíneoDocument19 pagesTecido SanguíneoBruno FelipeNo ratings yet
- O cérebro modifica-se quando aprendemosDocument6 pagesO cérebro modifica-se quando aprendemoscristina monizNo ratings yet
- SA 8 - ExlabDocument15 pagesSA 8 - ExlabChristhyanne Dos Santos CarreiroNo ratings yet
- Sebenta Das Aulas PDFDocument91 pagesSebenta Das Aulas PDFBruno PaulinoNo ratings yet
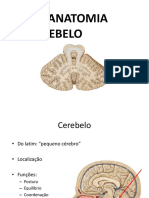
- Aula de Anatomia - Sistema Nervoso - CerebeloDocument5 pagesAula de Anatomia - Sistema Nervoso - CerebeloivairmatiasNo ratings yet
- Diencefalo PDFDocument12 pagesDiencefalo PDFjuvenal Enrique CoaquiraNo ratings yet
- Hipertireoidismo: conceito, etiologia, fisiopatologia e tratamentoDocument8 pagesHipertireoidismo: conceito, etiologia, fisiopatologia e tratamentokellenNo ratings yet
- Uretrocistografia Aula OBSDocument38 pagesUretrocistografia Aula OBSPauloCésarNo ratings yet
- Sistema digestório: funções, órgãos e digestãoDocument4 pagesSistema digestório: funções, órgãos e digestãoBrenda WilkeNo ratings yet
- Fisiologia Do Sistema DigestórioDocument3 pagesFisiologia Do Sistema DigestórioAnna Claara'No ratings yet
- Relatório Dissecção Do CoraçãoDocument9 pagesRelatório Dissecção Do Coraçãorui_silva19100% (6)
- Anatomia dos DentesDocument7 pagesAnatomia dos DentesJulya Moreira de Salvi AquinoNo ratings yet
- Sakura HemogramaDocument1 pageSakura HemogramalayssamaroNo ratings yet
- Massagem drenagem linfática manualDocument28 pagesMassagem drenagem linfática manualgustavogagiarNo ratings yet
- Anatomia Do Aparelho Reprodutor FemininoDocument57 pagesAnatomia Do Aparelho Reprodutor FemininoNinaNo ratings yet
- Músculos do corpo humano: principais características e funçõesDocument14 pagesMúsculos do corpo humano: principais características e funçõesRycher AguiarNo ratings yet
- Escultura Dental-AosdeiaDocument8 pagesEscultura Dental-AosdeiaJullya MagalhãesNo ratings yet
- Massagem DO-INDocument2 pagesMassagem DO-INmicaelrodrigo100% (4)
- Vascularização do membro superior: artérias, veias e linfáticosDocument53 pagesVascularização do membro superior: artérias, veias e linfáticosAna CoelhoNo ratings yet
- Orto SebentaDocument127 pagesOrto SebentaMiguel Caldeira PestanaNo ratings yet
- Apostila MIOLOGIADocument20 pagesApostila MIOLOGIACássia Depetris100% (2)
- Cronograma BFF I Vassouras 2023.1Document2 pagesCronograma BFF I Vassouras 2023.1Thiago MotaNo ratings yet
- 2º Ano Sistema Abo e Fator RHDocument7 pages2º Ano Sistema Abo e Fator RHRenato LimaNo ratings yet
- Estudo Dirigido - FluidosDocument2 pagesEstudo Dirigido - FluidosBeatriz EddineNo ratings yet
- Roteiro de Aula Prática RespiratórioDocument3 pagesRoteiro de Aula Prática Respiratórioneto fregatiNo ratings yet
- Estudos Dirigidos - Imuno Hematologia e Banco de SangueDocument15 pagesEstudos Dirigidos - Imuno Hematologia e Banco de SangueRaíssa AbreuNo ratings yet
- Telencéfalo 01 PDFDocument45 pagesTelencéfalo 01 PDFMarcelly FerreiraNo ratings yet
- Bate CoraçãoDocument1 pageBate CoraçãoRoberto Oliveira100% (1)
- Aula Anne LidaDocument50 pagesAula Anne LidaWillyan Oliveira da SilvaNo ratings yet