You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Can Musical Training Influence Brain ConnectivityDocument24 pagesCan Musical Training Influence Brain ConnectivityJohan NuñezNo ratings yet
- Cellulite: A Review of Its Physiology and TreatmentDocument6 pagesCellulite: A Review of Its Physiology and TreatmentShanna MitchellNo ratings yet
- Castellanos 2011 Alteration and ReorganizationDocument13 pagesCastellanos 2011 Alteration and ReorganizationSu AjaNo ratings yet
- Bokde, 2006.functional Connectivity of The Fusiform Gyrus During A Face-Matching Task in Subjects With MCI PDFDocument12 pagesBokde, 2006.functional Connectivity of The Fusiform Gyrus During A Face-Matching Task in Subjects With MCI PDFSu AjaNo ratings yet
- Niu2010.Cognitive Stimulation Therapy in The Treatment of Neuropsychiatric Symptoms in Alzheimer's Disease - A Randomized Controlled TrialDocument10 pagesNiu2010.Cognitive Stimulation Therapy in The Treatment of Neuropsychiatric Symptoms in Alzheimer's Disease - A Randomized Controlled TrialSu AjaNo ratings yet
- Rojas - Et - Al. - 2013.efficacy of A Cognitive Intervention Program in Patients With Mild Cognitive ImpairmentDocument7 pagesRojas - Et - Al. - 2013.efficacy of A Cognitive Intervention Program in Patients With Mild Cognitive ImpairmentSu AjaNo ratings yet
- Mills & Tamnes, 2014.methods and Considerations For Longitudinal Structural Brain Imaging Analysis Across DevelopmentDocument19 pagesMills & Tamnes, 2014.methods and Considerations For Longitudinal Structural Brain Imaging Analysis Across DevelopmentSu AjaNo ratings yet
- Mills & Tamnes, 2014.methods and Considerations For Longitudinal Structural Brain Imaging Analysis Across DevelopmentDocument10 pagesMills & Tamnes, 2014.methods and Considerations For Longitudinal Structural Brain Imaging Analysis Across DevelopmentSu AjaNo ratings yet
- Self Consciousness ScaleDocument11 pagesSelf Consciousness ScaleSu AjaNo ratings yet
- Van Os-2015-Cognitive Interventions in Older PDocument14 pagesVan Os-2015-Cognitive Interventions in Older PSu AjaNo ratings yet
- Bottiroli2008.Long-term Effects of Memory Training in The Elderly - A Longitudinal StudyDocument13 pagesBottiroli2008.Long-term Effects of Memory Training in The Elderly - A Longitudinal StudySu AjaNo ratings yet
- Styliadis-2015-Neuroplastic Effects of CombineDocument12 pagesStyliadis-2015-Neuroplastic Effects of CombineSu AjaNo ratings yet
- Papp, 2009.immediate and Delayed Effects of Co PDFDocument11 pagesPapp, 2009.immediate and Delayed Effects of Co PDFSu AjaNo ratings yet
- Mills & Tamnes, 2014.methods and Considerations For Longitudinal Structural Brain Imaging Analysis Across DevelopmentDocument19 pagesMills & Tamnes, 2014.methods and Considerations For Longitudinal Structural Brain Imaging Analysis Across DevelopmentSu AjaNo ratings yet
- ArticolDocument17 pagesArticolMinodora MilenaNo ratings yet
- Engvig, 2012.hippocampal Subfield Volumes Correlate With Memory Training Benefit in Subjective Memory Impairment PDFDocument7 pagesEngvig, 2012.hippocampal Subfield Volumes Correlate With Memory Training Benefit in Subjective Memory Impairment PDFSu AjaNo ratings yet
- Astle-2015-Cognitive Training Enhances IntrinsDocument7 pagesAstle-2015-Cognitive Training Enhances IntrinsSu AjaNo ratings yet
- Egvig, 2014..effects of Cognitive Training On GDocument13 pagesEgvig, 2014..effects of Cognitive Training On GSu AjaNo ratings yet
- Bahar Fuchs 2013 Cognitive Training and CognitDocument14 pagesBahar Fuchs 2013 Cognitive Training and CognitSu AjaNo ratings yet
- Egvig, 2014..effects of Cognitive Training On GDocument14 pagesEgvig, 2014..effects of Cognitive Training On GSu AjaNo ratings yet
- Belleville 2011 Training Related Brain PlasticDocument12 pagesBelleville 2011 Training Related Brain PlasticSu AjaNo ratings yet
- Nozawa-2015-Effects of Different Types of CognDocument18 pagesNozawa-2015-Effects of Different Types of CognSu AjaNo ratings yet
- Takeuchi-2010-Training of Working Memory ImpacDocument16 pagesTakeuchi-2010-Training of Working Memory ImpacSu AjaNo ratings yet
- Snarski-2011-The Effects of Behavioral Activat PDFDocument9 pagesSnarski-2011-The Effects of Behavioral Activat PDFSu AjaNo ratings yet
- NIH Public AccessDocument23 pagesNIH Public AccessSu AjaNo ratings yet
- Rodakowski 2015 Non Pharmacological InterventiDocument16 pagesRodakowski 2015 Non Pharmacological InterventiSu AjaNo ratings yet
- Bamidis, 2014.a Review of Physical and Cognitive Interventions in Aging.Document15 pagesBamidis, 2014.a Review of Physical and Cognitive Interventions in Aging.Su AjaNo ratings yet
- Teipel, 2012.anatomical MRI and DTI in The DiagDocument16 pagesTeipel, 2012.anatomical MRI and DTI in The DiagSu AjaNo ratings yet
- Sala Llonch 2015 Reorganization of Brain NetwoDocument11 pagesSala Llonch 2015 Reorganization of Brain NetwoSu AjaNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Pengaruh Media Sosial Instagram Dan Whatsapp Terhadap Pembentukan Budaya "Alone Together"Document12 pagesPengaruh Media Sosial Instagram Dan Whatsapp Terhadap Pembentukan Budaya "Alone Together"Muhammad Farhan SyahNo ratings yet
- Aula PulpDocument64 pagesAula PulpFelipe Rech BrunettaNo ratings yet
- Paul Martin Postal - Three Investigations of ExtractionDocument229 pagesPaul Martin Postal - Three Investigations of ExtractionpalingenesiaNo ratings yet
- Chapter 7 QuestionsDocument6 pagesChapter 7 QuestionsArnav Pareek0% (2)
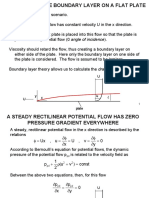
- Boundary Layer Flat PlateDocument27 pagesBoundary Layer Flat PlateGohar KhokharNo ratings yet
- Objective Mapping and Kriging: 5.1 Contouring and Gridding ConceptsDocument24 pagesObjective Mapping and Kriging: 5.1 Contouring and Gridding Conceptsforscribd1981No ratings yet
- ELE Final MCQDocument141 pagesELE Final MCQSanket KarandeNo ratings yet
- Consumer's EquilibriumDocument4 pagesConsumer's EquilibriumMahendra ChhetriNo ratings yet
- Problems and Solutions For Complex AnalysisDocument259 pagesProblems and Solutions For Complex AnalysisMete Torun91% (34)
- Atkinson 2003Document64 pagesAtkinson 2003Akshay29No ratings yet
- Chapter 2/number 61/problem Set 2.3 Ferris Wheel in 1897, A Ferris Wheel Was Built in Vienna That Still Stands Today. It Is Named TheDocument3 pagesChapter 2/number 61/problem Set 2.3 Ferris Wheel in 1897, A Ferris Wheel Was Built in Vienna That Still Stands Today. It Is Named TheINKA VERANDERA NUGRAHANo ratings yet
- m2 CH 3 Binomial Theorem Supp ExDocument1 pagem2 CH 3 Binomial Theorem Supp Exapi-303501215No ratings yet
- Concept of StrategyDocument3 pagesConcept of StrategyvikasmishraelexNo ratings yet
- Sky ServerDocument30 pagesSky ServerMd RashidunnabiNo ratings yet
- Namma Kalvi 12th Accountancy Unit 4 Sura English Medium Guide PDFDocument10 pagesNamma Kalvi 12th Accountancy Unit 4 Sura English Medium Guide PDFAakaash C.K.No ratings yet
- Multinomial Logistic Regression Models: Newsom Psy 525/625 Categorical Data Analysis, Spring 2021 1Document5 pagesMultinomial Logistic Regression Models: Newsom Psy 525/625 Categorical Data Analysis, Spring 2021 1Deo TuremeNo ratings yet
- Trust-In Machine Learning ModelsDocument11 pagesTrust-In Machine Learning Modelssmartin1970No ratings yet
- Reviewed Risk Theory NotesDocument112 pagesReviewed Risk Theory Notesajami malalaNo ratings yet
- Project Management: CrashingDocument22 pagesProject Management: CrashingRushi JivaniNo ratings yet
- Lecture # 23: Segmentation Cont.: Muhammad Rzi AbbasDocument12 pagesLecture # 23: Segmentation Cont.: Muhammad Rzi AbbasMA KhanNo ratings yet
- Javacc FaqDocument66 pagesJavacc FaqFrederick Bejarano SanchezNo ratings yet
- Adobe Scan May 23, 2022Document3 pagesAdobe Scan May 23, 2022Anagh ShuklaNo ratings yet
- Body in White Architecture For An Electric Vehicle ConceptDocument54 pagesBody in White Architecture For An Electric Vehicle ConceptJosue Enriquez Eguiguren100% (1)
- GplayDocument9 pagesGplayΣατανάς ΣατανάςNo ratings yet
- Complex NumbersDocument87 pagesComplex NumbersPriya DudetteNo ratings yet
- ABACUS Features USDocument2 pagesABACUS Features USSuzanne StewartNo ratings yet
- Special Relativity and Minkowski SpacesDocument13 pagesSpecial Relativity and Minkowski Spacesgraviton6No ratings yet
- Basic Math - 2011 PDFDocument6 pagesBasic Math - 2011 PDFLuqman Abdirazaq TukeNo ratings yet
- Lista Ejercicios 3 PPEDocument25 pagesLista Ejercicios 3 PPEKarla HermorNo ratings yet
- Instrumentation & Control HandoutDocument95 pagesInstrumentation & Control HandoutPandimadevi SelvakumarNo ratings yet