You might also like
- Feldenkrais Reloj PelvicoDocument5 pagesFeldenkrais Reloj PelvicoAnonymous Dv72G9joNo ratings yet
- Guia Regalo Recetas Caseras Variedades Nice PDFDocument18 pagesGuia Regalo Recetas Caseras Variedades Nice PDFmarioli90% (10)
- Ponencia para Esge-Fiag Julio 2018Document30 pagesPonencia para Esge-Fiag Julio 2018José LuisNo ratings yet
- Ejercicios de Matematica Financiera 2Document66 pagesEjercicios de Matematica Financiera 2Nansy Grijalva54% (26)
- Practica Calificada 1 (Ensayo) - Grupo 8Document3 pagesPractica Calificada 1 (Ensayo) - Grupo 8Amador Raymundo100% (1)
- Martensita RevenidaDocument3 pagesMartensita RevenidaIsmael MelendezNo ratings yet
- Hipertension OcularDocument2 pagesHipertension OcularDr. Javier Antonio Toledo LimónNo ratings yet
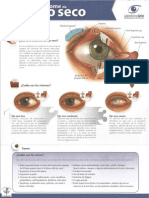
- Sindrome Ojo SecoDocument2 pagesSindrome Ojo SecoDr. Javier Antonio Toledo LimónNo ratings yet
- Diabetes Tipo 2-Algoritmo de TratamientoDocument6 pagesDiabetes Tipo 2-Algoritmo de TratamientoDr. Javier Antonio Toledo LimónNo ratings yet
- ProstataDocument48 pagesProstataDr. Javier Antonio Toledo LimónNo ratings yet
- GlaucomaDocument2 pagesGlaucomaDr. Javier Antonio Toledo LimónNo ratings yet
- RiñonDocument2 pagesRiñonDr. Javier Antonio Toledo LimónNo ratings yet
- Degeneracion MacularDocument2 pagesDegeneracion MacularDr. Javier Antonio Toledo LimónNo ratings yet
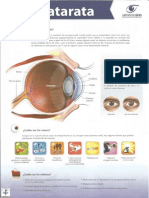
- CatarataDocument2 pagesCatarataDr. Javier Antonio Toledo LimónNo ratings yet
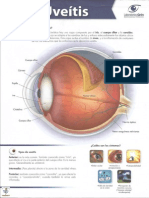
- UveitisDocument2 pagesUveitisDr. Javier Antonio Toledo LimónNo ratings yet
- Estudios Radiograficos PediatriaDocument3 pagesEstudios Radiograficos PediatriaDr. Javier Antonio Toledo LimónNo ratings yet
- Tratamiento Hipertension Arterial Sistemica Caso ClinicoDocument15 pagesTratamiento Hipertension Arterial Sistemica Caso ClinicoDr. Javier Antonio Toledo Limón100% (1)
- Traumatismos DeporteDocument12 pagesTraumatismos DeporteDr. Javier Antonio Toledo LimónNo ratings yet
- Reflujo GastroesofágicoDocument5 pagesReflujo GastroesofágicoDr. Javier Antonio Toledo LimónNo ratings yet
- Mercadotecnia Practica MedicaDocument11 pagesMercadotecnia Practica MedicaDr. Javier Antonio Toledo LimónNo ratings yet
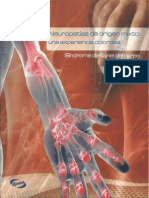
- Neuropatia Nervio MusculocutaneoDocument7 pagesNeuropatia Nervio MusculocutaneoDr. Javier Antonio Toledo LimónNo ratings yet
- Trastornos Venosos PerifericosDocument11 pagesTrastornos Venosos PerifericosDr. Javier Antonio Toledo LimónNo ratings yet
- Terapia Reemplazo HormonalDocument2 pagesTerapia Reemplazo HormonalDr. Javier Antonio Toledo LimónNo ratings yet
- Sindrome Tunel TarsianoDocument5 pagesSindrome Tunel TarsianoDr. Javier Antonio Toledo LimónNo ratings yet
- Sindrome Tunel CarpoDocument7 pagesSindrome Tunel CarpoDr. Javier Antonio Toledo LimónNo ratings yet
- Mi Vida Con Hipertension ArterialDocument6 pagesMi Vida Con Hipertension ArterialDr. Javier Antonio Toledo LimónNo ratings yet
- CorticoterapiaDocument25 pagesCorticoterapiaDr. Javier Antonio Toledo LimónNo ratings yet
- Laboratoriales Examenes DiabetesDocument20 pagesLaboratoriales Examenes DiabetesDr. Javier Antonio Toledo LimónNo ratings yet
- GlaucomaDocument2 pagesGlaucomaDr. Javier Antonio Toledo LimónNo ratings yet
- Infecciones Genitales No Complicadas Preguntas RespuestasDocument9 pagesInfecciones Genitales No Complicadas Preguntas RespuestasDr. Javier Antonio Toledo LimónNo ratings yet
- Caso Clinico Radiculopatia LumbarDocument7 pagesCaso Clinico Radiculopatia LumbarDr. Javier Antonio Toledo LimónNo ratings yet
- Estreñimiento BebeDocument6 pagesEstreñimiento BebeDr. Javier Antonio Toledo LimónNo ratings yet
- Hogar Seguro AlzheimerDocument6 pagesHogar Seguro AlzheimerDr. Javier Antonio Toledo LimónNo ratings yet
- Hiperglicemia Hipoglucemia Consecuencias EjerciciosDocument11 pagesHiperglicemia Hipoglucemia Consecuencias EjerciciosDr. Javier Antonio Toledo LimónNo ratings yet
- Caso Clinico Sindrome Alergia Multiples MedicamentosDocument5 pagesCaso Clinico Sindrome Alergia Multiples MedicamentosDr. Javier Antonio Toledo LimónNo ratings yet
- Comic Hiperplasia Prostatica BenignaDocument15 pagesComic Hiperplasia Prostatica BenignaDr. Javier Antonio Toledo LimónNo ratings yet
- C 05 Cuencas HidrográficasDocument24 pagesC 05 Cuencas HidrográficasSolNo ratings yet
- Conclusiones ClifDocument3 pagesConclusiones ClifJoselyn Rosario SincheNo ratings yet
- Gubler Pascual Rojas SanchezDocument54 pagesGubler Pascual Rojas SanchezJazmin Alicia Pantoja RojasNo ratings yet
- Ensayo - Evaluación NeuropsicológicaDocument8 pagesEnsayo - Evaluación NeuropsicológicaLina Cervantes100% (1)
- Síntesis de (Ti〖 (Urea) 〗 - 6) I - 3 PrecasiDocument3 pagesSíntesis de (Ti〖 (Urea) 〗 - 6) I - 3 PrecasiLeoin Garcia ValleNo ratings yet
- In vitro propagation of Agave cocui through axillary budsDocument7 pagesIn vitro propagation of Agave cocui through axillary budsAnabel HernándezNo ratings yet
- Producto MilagrosDocument5 pagesProducto MilagrosGerardo MvNo ratings yet
- Formato de Evaluación de Cosecha ArándanoDocument1 pageFormato de Evaluación de Cosecha Arándanoluis perezNo ratings yet
- 230 Difteria 2019Document5 pages230 Difteria 2019andresguastarNo ratings yet
- Manualcap 3Document24 pagesManualcap 3JORGE STIVENS MONCADA PERLAZANo ratings yet
- Tarea 3 Psicologia GeneralDocument6 pagesTarea 3 Psicologia GeneralalfonsinaNo ratings yet
- Cambio ClimaticoDocument8 pagesCambio ClimaticoMh LizNo ratings yet
- Actividades Aprendizaje - Agenda No. 4Document3 pagesActividades Aprendizaje - Agenda No. 4brayan Cortez0% (1)
- PROC-005 PROCEDIMIENTOS DE ALMACENAMIENTO DE PRODUCTOS FARMACÉUTICOS, DISPOSITIVOS MÉDICOS Y PRODUCTOS SANITARIOS.. Rev 0Document3 pagesPROC-005 PROCEDIMIENTOS DE ALMACENAMIENTO DE PRODUCTOS FARMACÉUTICOS, DISPOSITIVOS MÉDICOS Y PRODUCTOS SANITARIOS.. Rev 0Lio MarinNo ratings yet
- Capitulo Iii PDFDocument46 pagesCapitulo Iii PDFedwinNo ratings yet
- Práctica N°2 sobre materiales poliméricosDocument21 pagesPráctica N°2 sobre materiales poliméricosRichard Ventura AndacabaNo ratings yet
- Guía #2Document8 pagesGuía #2Shelsy BuzónNo ratings yet
- MITT Manual de Usuario 125 GPDocument64 pagesMITT Manual de Usuario 125 GPAlberto Núñez MéndezNo ratings yet
- Principales Plagas de QuercusDocument12 pagesPrincipales Plagas de QuercusJuan BucheliNo ratings yet
- COLIFORMESDocument4 pagesCOLIFORMESFaNi Saucedo C50% (2)
- Instalaciones SanitariasDocument16 pagesInstalaciones SanitariashansNo ratings yet
- El Sentido de La VidaDocument8 pagesEl Sentido de La VidaGomez calderon Karol Marisol.No ratings yet
- Planificación docente de Castellano en escuela agropecuariaDocument2 pagesPlanificación docente de Castellano en escuela agropecuariayenny vivas0% (1)
- Suicidio y Depresión en Adolescentes: Una Revisión de La LiteraturaDocument12 pagesSuicidio y Depresión en Adolescentes: Una Revisión de La LiteraturaFraga DavidNo ratings yet