Professional Documents
Culture Documents
Genetica Bacteriana
Uploaded by
Rayito de SolOriginal Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Genetica Bacteriana
Uploaded by
Rayito de SolCopyright:
Available Formats
CAPTULO 4 GENTICA BACTERIANA
Germn Hermosilla D.
INTRODUCCIN La informacin gentica en bacterias est codificada en la molcula de cido desoxirribonucleco, ADN, cuya propiedad de replicacin es esencial para la herencia de todos los rasgos genticamente determinados (genotipo) y que en su conjunto definen las caractersticas observables del microorganismo (fenotipo). Este captulo entregar al alumno los elemento bsicos relacionados con la organizacin de la informacin gentica en las bacterias, su relacin con rasgos fenotpicos de inters mdico (factores de virulencia y resistencia a antibiticos), as como, los mecanismos por los cuales estos organismo procariticos pueden adquirir variabilidad gentica. NATURALEZA DEL MATERIAL HEREDITARIO EN BACTERIAS La molcula de ADN, corresponde a un polmero de cido nucleico, cuya unidad es el nucletido. Cada nucletido posee un grupo fosfato, una azcar desoxirribosa y una base nitrogenada purina o pirimidina. Las purinas son adenina (A) y guanina (G), mientras que las pirimidinas corresponden a timina (T) y citocina (C). Los nucletidos se unen uno a uno a travs de un enlace covalente llamado unin fosfodiester, en la que un grupo 3-hidroxilo de un nucletido se une al grupo 5-fosfato del nucletido adyacente. Esta unin fosfodiester asimtrica define la polaridad de la cadena polinucleotdica. El ADN es una molcula de doble hebra, con forma helicoidal, donde las dos hebras en la hlice son antiparalelas. La doble hlice se encuentra estabilizada por uniones dbiles del tipo puente de hidrgeno, entre una base purina y otra pirimidina, en hebras opuestas. En cada posicin del ADN, la adenina se aparea por dos puentes de hidrgeno a timina en la hebra opuesta, mientras que guanina se aparea por tres puentes de hidrgeno a una citocina. Por lo tanto, las dos hebras de la doble hlice son complementarias. Debido a esta complementaridad, la doble hebra de ADN contiene cantidades equimolares de purinas (A+G) y pirimidinas (T+C), con igual cantidad de A y T, como G y C. Sin embargo, la fraccin molar de G+C en el ADN puede variar entre las diferentes especies de bacterias. El ADN posee dos funciones esenciales en los microorganismos, la replicacin y la expresin. Replicacin: Durante la replicacin, cada hebra de la doble hlice sirve como templado para la sntesis de una hebra complementaria. Cada nueva molcula de doble hebra de ADN debe contener, por lo tanto, una hebra polinucleotdica vieja y una hebra recientemente sintetizada.
27
28 Biologa de los microorganismos
Este tipo de replicacin del ADN se denomina semiconservativa. Por otro lado, la replicacin del ADN cromosomal en bacterias comienza en un sitio especfico llamado origen y se realiza bidireccionalmente hasta que el proceso se completa. De este modo, cuando una bacteria se divide por fisin binaria, luego de haber completado la replicacin de su ADN, cada clula hija hereda un genoma replicado. Expresin: La informacin gentica codificada en el ADN se expresa a travs de la sntesis de otro cido nucleico, el ARN y protenas. La sntesis de ARN, dependiente del ADN, se denomina transcripcin, mientras que la sntesis de las protenas a partir del ARN se llama traduccin. La direccin del flujo de informacin entre estas molculas esta definido por el Dogma de la Biologa Molecular (Figura 4-1).
ADN
ARN
retrovirus
protena
ARN virus
Figura 4-1: Dogma central de la biologa molecular.
La expresin gnica en bacterias est regulada. De esta forma el microorganismo se adapta rpidamente a la variacin de concentracin de nutrientes presentes en el ambiente que los rodea. Para ello, las bacterias poseen genes relacionados, agrupados estructuralmente en operones. Genes relacionados pueden ser aquellos que codifican para enzimas que participan en una va metablica comn. Un opern consiste de un solo promotor (secuencia reguladora de la transcripcin), genes agrupados (cistrones) y de un terminador de la transcripcin. De esta forma, los genes son transcritos coordinadamente en un ARN mensajero policistrnico, que luego ser traducido a protenas. Las bacterias, regulan la expresin gnica a nivel de la transcripcin y la traduccin. A pesar de su importancia, ambos procesos no sern revisados en este captulo, dado que lo concerniente a regulacin de la expresin fue abordado en el captulo de Fisiologa Bacteriana. Se debe mencionar que las bacterias poseen tambin niveles de regulacin superior al opern. Estos constituyen el Sistema de Regulacin Global, el que est constituido por los regulones: un conjunto de operones asociados a una va, funcin o proceso comn, bajo el control de la misma protena reguladora; los modulones: un conjunto de operones relacionados a vas o funciones mltiples, que responden a una protena reguladora comn; y los estimulones: un conjunto de operones, regulones o modulones, que responden frente a una seal ambiental comn.
Gentica bacteriana 29
ORGANIZACIN GENTICA EN BACTERIAS La informacin heredable en bacterias est organizada en dos elementos genticos: el cromosoide o genoma bacteriano y elementos extracromosmicos, como los plsmidos y bacterifagos. Estos elementos, replicables como unidades genticas discretas, son llamados genricamente replicones. Cromosoide bacteriano: El tamao del genoma bacteriano vara entre 0.4x109 a 8.6x109 dalton. Los cromosoides ms pequeos se encuentran en las bacterias parsitos obligados, tales como micoplasmas, mientras que los de mayor tamao estn en bacterias capaces de una diferenciacin compleja, tal como Myxococcus. La cantidad de ADN en el genoma determina la cantidad mxima de informacin que puede codificar. La mayora de las bacterias presentan un genoma haploide, de una molcula simple de ADN, con topologa circular. Sin embargo, la bacteria Gram (-) Borrelia burgdorferi, posee un cromosoide lineal, mientras que Agrobacterium tumefaciens cuenta con un cromosoide lineal y otro circular. Por otro lado, continuando con las excepciones, existen bacterias cuyos genomas son diploides (Leptospira interrogans) e incluso triploides (Burkholderia cepacia). El cromosoide de Escherichia coli es de 3x109 Da, 4500 kilobases (kb), lo que supone una longitud de 1.3 mm, aproximadamente 1000 veces el dimetro de la clula. Esto se explica debido a que el ADN del cromosoide est superenrollado y altamente empaquetado en el nucleoide bacteriano. Se estima que el genoma de E. coli codifica para alrededor de 4000 genes, que definen las caractersticas fenotpicas de esta bacteria. El tiempo requerido para la replicacin completa de este cromosoide es de alrededor de 40 min.
Plsmidos: Los plsmidos son replicones que se mantienen en las bacterias como elementos genticos extracromosomales y discretos (Figura 4-2). En general son de menor tamao que el cromosoide bacteriano entre 5 a varias decenas de kb, aunque plsmidos tan grandes como 2 Mb pueden encontrarse en bacterias. Los plsmidos normalmente codifican para rasgos fenotpicos no esenciales para la viabilidad de la bacteria y se replican en forma independiente del cromosoide. Casi todos los plsmidos son molculas de doble hebra de ADN y circulares, aunque tambin se han reportado plsmidos lineales en Borrelia y Streptomyces. Dos plsmidos estrechamente relacionados o idnticos no pueden ser mantenidos en forma estable dentro de un mismo hospedero bacteriano, por lo tanto son incompatibles. mientras que plsmidos no relacionados pueden coexistir, dado que pertenecen a grupos de incompatibilidad diferentes. La incompatibilidad queda establecida por la competencia de los factores (protenas del hospedero) necesarios para la replicacin plasmidial.
30 Biologa de los microorganismos
PBR322
ampR 4.4 kb
tetR
ori
Figura 4-2: Representacin de un plsmido tpico. ori, origen de replicacin; ampR, gen de resistencia a ampicilina; tetR, gen de resistencia a tetraciclina. Se destacan sitios blancos de enzimas de restriccin.
De acuerdo a las caractersticas que pueden conferir a las bacterias que los portan, los plsmidos pueden ser clasificado como: Plsmidos R: Plsmidos col: Plsmidos metablicos: Aquellos que codifican para resistencia a antibiticos. Los que expresan factores que actan como bacteriocinas. Los que portan los genes necesarios para el metabolismos de molculas complejas. Plsmidos de virulencia: Plsmidos que codifican para factores de virulencia. Plsmidos conjugativos: Aquellos que tienen la informacin necesaria para promover su propia transferencia a otra bacteria receptora.
El nmero promedio de molculas de un plsmido particular por bacteria se denomina nmero de copia. Plsmidos mayores a 40 kb son frecuentemente conjugativos y tienen un bajo nmero de copia (1 a 10 por bacteria). Estos codifican para todas las funciones requeridas para su replicacin y particin en las clulas hijas. En cambio, plsmidos menores que 7.5 kb, normalmente no son conjugativos y tienen una alto nmero de copias (entre 20 a 50 por bacteria) Muchos plsmidos controlan importantes propiedades patognicas de las bacterias patognicas, como la resistencia a los antibiticos, la produccin de toxinas y la sntesis de estructuras celulares de superficie requeridas para la adherencia o colonizacin e invasin. Por
Gentica bacteriana 31
ejemplo, toxinas representativas, codificados por plsmidos, incluyen las enterotoxinas termo lbil y termo estable de E. coli, la toxina tetnica de Clostridium tetani y la toxina exfoliativa de Staphylococcus aureus. Por otro lado, un plsmido virulento ha sido reportado en Yersinia, en donde el elemento extracromosmico codifica para las protenas YOps requeridas en la invasin de esta bacteria.
Bacterifagos: Los bacterifagos (virus bacterianos) son agentes infecciosos que se replican como un parsito obligado en bacterias. El fago extracelular es metablicamente inerte y consiste principalmente de protenas mas cido nucleico (ADN o ARN). Las protenas del fago protegen su material gentico (cpside). El genoma del fago puede variar de 2 a 200 kb por hebra de ADN y su genoma puede ser una molcula de simple hebra o doble hebra. Como ocurre en los plsmidos, los fagos codifican para las funciones necesarias para su replicacin en bacterias, pero adems, protenas de la cpside y protenas no estructurales requeridas para el ensamblaje del fago. Existen varios tipos morfolgicos de fagos, los hay polihdricos, filamentosos y complejos. Estos ltimos, tienen cabezas polidricas a las cuales se une una cola y a veces otros apndices. El paso inicial de la infeccin de un fago es la adsorcin de ste a receptores especficos en la superficie del hospedero bacteriano susceptible. La cpside permanece en la superficie celular, y el genoma ARN o ADN entra en la clula blanco (penetracin). El genoma del fago es replicado para producir nuevas copias, y las protenas fago-especficas son expresadas. En la mayora de los fagos, el ensamblaje de la progenie ocurre en el citoplasma y la liberacin de esta ocurre por lisis celular. Sin embargo, los fagos filamentosos son formados en la envoltura celular y liberados con la muerte de la bacteria. El nmero promedio de partculas vricas producida dentro de cada hospedero es caracterstico de cada fago y frecuentemente vara entre 50 y varios cientos. Los fagos son clasificados en dos grandes grupos: Fagos virulentos y fagos temperados. El ciclo de los fagos virulento en bacterias susceptibles destruye la clula del hospedero. En cambio, los fagos temperados pueden seguir un ciclo ltico o lisognico. El ciclo ltico de los bacterifagos temperados y virulentos es similar, terminando con la produccin de la progenie vrica y la muerte del hospedero bacteriano. El ciclo lisognico es un tipo especfico de infeccin viral latente, en la cual el genoma del fago se replica como un profago dentro de la clula bacteriana. En la mayora de las bacterias lisognicas los genes requeridos para el ciclo ltico no son expresados. El estado fsico del profago no es idntico en todos los fagos. Por ejemplo, el profago del bacterifago 1 en E. coli se encuentra integrado al cromosoide bacteriano, en un sitio especfico y se replica como parte de l, mientras que el profago P1 en E. coli se replica como un plsmido extracromosomal. En bacterias lisognicas, el profago puede activarse entrando al ciclo ltico. El estado ltico puede ser inducido en forma sincrnica cuando la bacteria lisognica es tratada con agentes que daan el ADN (Luz ultravioleta o mitomicina C).
32 Biologa de los microorganismos
Algunos fagos temperados contienen genes para caracterstica bacterianas que no estn relacionados a ciclo ltico o estado lisognico. La expresin de tales genes se denomina conversin del fago o conversin lisognica. Ejemplos de conversin lisognica importantes en la virulencia bacteriana incluyen la produccin de la toxina de la difteria en Corynebacterium diphtheriae, la toxina eritrognica en Streptococcus pyogenes, toxina botulnica en Clostridium botulinum y las toxinas tipo-Shiga en E. coli. Los genes que codifican para estas toxinas estn presentes en el genoma del fago temperado. Por otro lado, la especificidad de los antgenos O en Salmonella puede estar tambin controlado por la conversin lisognica.
OTROS ELEMENTOS GENTICOS EN BACTERIAS Adems de los plsmidos y bacterifagos, existen otros elementos genticos presentes en bacterias. A diferencia de los primeros estos elementos no son capaces de replicarse en forma autnoma. Su mantencin en la bacteria depende de otro replicn, como el cromosoide bacteriano, plsmido o bacterifago. Estos elementos son el transposn y los integrones. Transposn: Los transposones son segmentos de ADN, los cuales pueden moverse desde un sitio en una molcula de ADN a otro sitio blanco en la misma o una diferente la molcula de ADN. El proceso se denomina transposicin y ocurre por un mecanismo independiente de la recombinacin generalizada. Los transposones son elementos genticos importantes ya que pueden causar mutaciones, mediar rearreglos cromosmicos, portar regiones de ADN de homologa gentica, y adquirir nuevos genes, los que pueden ser diseminados en la poblacin bacteriana. La insercin de un transposn normalmente interrumpe la secuencia lineal de un gen y los inactiva. La mayora de los transposones comparten rasgos en comn. Todo transposn codifica las funciones necesarias para su transposicin, incluyendo una enzima transposasa que interacta con secuencias especficas en los extremos del transposn. Durante el evento de transposicin, una corta secuencia de ADN es duplicada, y el transposn se inserta entre las secuencias repetidas directas. El tamao de esta corta secuencia es variable y es caracterstica de cada transposn. Algunos transposones pueden insertarse en casi cualquier secuencia blanco, mientras que otros son altamente especficos. Se reconocen dos tipos de transposicin. La excisin del transposn desde un sitio, para luego insertarse en otro sitio blanco se denomina transposicin no-replicativa. Si el transposn en un sitio dador es replicado y una copia se inserta en el sitio blanco, el proceso se llama transposicin replicativa.
Gentica bacteriana 33
Los transposones en bacterias pueden ser separados en tres mayores clases (Figura 4-3).
1) Secuencias de Insercin (IS): Estos elementos son simples en estructura y codifican slo para las funciones necesarias para la transposicin. Su tamao vara entre 780 a 1500 pb, tienen repetidos invertidos cortos en sus extremos (15-25 pb). El ADN entre los repetidos invertidos terminales contiene un gen de la transposasa, excepcionalmente dos. Los transposones compuestos varan en longitud de 2000 pb a 40000 bp y tienen secuencias de insercin en sus extremos, normalmente como repetidos invertidos. En bacterias de inters mdico, los transposones compuestos pueden portar genes que determinan la adhesin, produccin de toxinas y otros factores de virulencia, as como genes de resistencia a uno o ms antibiticos. Por ejemplo, los transposones Tn5 y Tn10, determinan la resistencia a kanamicina y tetraciclina, respectivamente. 2) Transposones de la familia TnA: Estos transposones poseen repetidos terminales invertidos de mayor tamao que los transposones compuestos (35 a 40 pb) y adems carecen de secuencias de insercin terminales. Todos los miembros de este tipo de transposn codifica para la transposasa y otra enzima la resolvasa. Dos transposones de esta clase son Tn3 (resistencia a ampicilina) y Tn1000, encontrados en el plsmido conjugativo F. 3) Bacteriofago Mu y fagos temperados relacionados: El genoma completo de este elemento funciona como un transposn. La replicacin del ADN del fago, durante el crecimiento vegetativo, ocurre por transposicin replicativa. La integracin del profago puede ocurrir en diferentes sitios del cromosoide bacteriano y frecuentemente causa mutaciones. Por esta razn Mu y los fagos relacionados son a veces llamado fagos mutadores.
Existe una cuarta clase de transposones, recientemente descubierta en bacterias Gram (+). Son denominados transposones conjugativos. El transposn Tn917 pertenece a esta clase. Estos transposones son totalmente diferentes de los descritos anteriormente. Estos no generan duplicacin de las secuencias blancos en las cuales se inserta. Adems, los transposones conjugativos, de forma similar a los plsmidos conjugativos, tienen la informacin necesaria para ser transferido de una bacteria a otra mediante el proceso de conjugacin. Sin embargo, a diferencia de la conjugacin realizada por los plsmidos conjugativos, la conjugacin por transposones conjugativos no necesita que las bacterias a conjugar estn relacionadas estrechamente. Estos transposones son los principales responsables de la diseminacin de resistencia en bacterias Gram (+).
34 Biologa de los microorganismos
Secuencia de Insercin
Transposasa
1
Transposn compuesto (Tn5) IS50L Transposasa Resistencia a kanamicina Km IS50R Transposasa
Familia Tn A (Tn3)
Ap Transposasa Resolvasa Resistencia a ampicilina
Fago
Genes Fago (cabeza/cola) Transposasa
Figura 4-3:
Clases de transposones representativos, integrados en el genoma bacteriano. El rectngulo con barras oblicuas representa la repetido terminales invertidos. Las puntas de flecha en blanco corresponde al sitio de insercin del transposn, repetido en la misma orientacin.
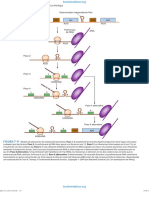
Integrones: Estos elementos genticos son importantes en la evolucin de los transposones. La primera indicacin de su importancia provienen de la comparacin de los miembros de la familia Tn21 (Figura 4-4). Los integrones tienen en comn repetidos invertidos en sus extremos y una gen que codifica para la enzima integrasa. Esta enzima media la insercin de genes de resistencia a antibiticos (cassette gnicos) en una secuencia especfica GTT. Todos los integrones poseen un promotor fuerte, denominado sitio P, que controla la expresin de los genes de resistencia integrados. En la Figura 4-4, se muestran tres integrones, los cuales son similares, excepto que ellos tienen diferentes genes de resistencia a antibiticos, insertados exactamente en el mismo sitio P. El transposn In0 no contiene ningn gen de resistencia a antibitico en el sitio P, mientras que In3 tiene un gen para resistencia a trimethoprim en este sitio. In2, el cual es realmente un transposn insertado dentro del Tn21, tiene un gen de resistencia a streptomicina y espectinomicina en el sitio P.
Gentica bacteriana 35
GTT
GTT
59 pb
GTT
59 pb
Figura 4-4:
Integrones de la familia Tn21. IR, representa una secuencia repetida invertida de 25 pb; int, gen de la integrasa; sul1, resistencia a sulfonamida; dhfr, resistencia a trimetoprim; aadA , resistencia a estreptomicina y espectinomicina; GTT, Sitio de insercin del casset gnico de resistencia; 59 pb, elemento de 59 pb involucrado en los eventos de recombinacin mediada por la integrasa; P(P2), promotor fuerte.
VARIACIN GENTICA EN BACTERIAS A pesar que las bacterias se multiplican clonalmente, por fisin binaria, en las poblaciones bacterianas la existencia de variabilidad gentica es un hecho irrefutable. Existen bsicamente dos mecanismos por los cuales las bacterias adquieren variabilidad: la mutacin y la transferencia gentica horizontal.
Mutaciones: Las mutaciones son cambios heredables en el genoma. stas pueden ocurrir en forma espontnea durante la replicacin, o ser inducidas por agentes mutagnicos qumicos (etilmetanosulfanato) o fsicos (luz ultravioleta). Las mutaciones pueden causar cambios en las caractersticas fenotpicas de una bacteria y constituyen la fuente primordial de variabilidad gentica en todos los seres vivos. Para que una mutacin se establezca en trminos poblacionales, debe conferir una mejor adaptacin de la bacteria a su entorno, de tal forma que ocurra seleccin positiva del nuevo clon. Las mutaciones son clasificadas sobre la base de los cambios estructurales que pueden ocurrir en el ADN. Algunas mutaciones son localizadas dentro de cortos segmentos de ADN (substituciones nucleotdicas, microdeleciones y microinserciones). Otras, en cambio,
36 Biologa de los microorganismos
involucran grandes regiones del ADN e incluyen las deleciones, inserciones o rearreglos cromosmicos.
Transferencia horizontal de informacin gentica: La transferencia horizontal de informacin gentica provee a las bacterias de un mecanismo de sexualidad. La interaccin gentica entre los microorganismos permite que sus genomas evolucionen mucho ms rpidamente que por el proceso de mutacin y seleccin nicamente. El intercambio de informacin gentica entre bacterias ha tenido gran impacto en biomedicina, a travs de la emergencia y diseminacin de plsmidos de resistencia a antibiticos, variacin de fase flagelar en Salmonella y variacin antignica de antgenos de superficie en Neisseria y Borrelia. La transferencia horizontal en bacterias involucra el intercambio de informacin gentica desde una clula dadora a otra clula recipiente. La transformacin, transduccin y conjugacin son procesos sexuales que utilizan diferentes mecanismos para introducir ADN donador en una bacteria recipiente. Debido a que el ADN donador no puede persistir en la bacteria recipiente, a menos que sea parte de un replicn (plsmido, bacterifago), la recombinacin entre los genomas dador y recipiente es requerida frecuentemente para producir progenie hbrida estable.
Transformacin: En la transformacin, las molculas de ADN provenientes desde la bacteria donadora son captadas directamente desde el ambiente extracelular por la bacteria recipiente. La recombinacin ocurre entre las molculas simples del ADN transformante y el cromosoide bacteriano. El ADN transformante debe ser al menos de 500 pb y corresponde a fragmentos pequeos del cromosoma bacteriano. La transformacin fue descubierta primero en Streptococcus pneumoniae, pero puede darse en otros gneros de bacterias como Haemophilus, Neisseria, Bacillus y Staphylococcus. La habilidad de la bacteria para captar ADN extracelular y llegar a ser transformada se denomina competencia y depende del estado fisiolgico de la bacteria. En algunas bacterias la captacin de ADN depende de la presencia de secuencias especficas en el ADN transformante (Haemophilus y Neisseria), mientras que en otros gneros no se requiere una secuencia especfica (Streptococcus pneumoniae). La bacteria competente puede adems captar ADN intacto, como el presente en bacterifago (transfeccin) o plsmidos, los cuales pueden replicarse como elementos extracromosmicos en la bacteria recipiente.
Transduccin: En la transduccin, los bacterifagos funcionan como vectores para introducir ADN desde una bacteria dadora en una bacteria recipiente por infeccin. Para algunos fagos, llamados fagos de transduccin generalizada, una pequea fraccin de los fagos producidos durante el ciclo ltico son aberrantes y contienen fragmentos aleatorios del cromosoide bacteriano, en lugar del ADN del fago. Cada fago transductante individual puede transportar sets diferentes de genes estrechamente ligados, que representan un pequeo segmento del genoma bacteriano. La transduccin mediada por una poblacin de tales fagos es llamada
Gentica bacteriana 37
transduccin generalizada, debido a que cualquier regin del genoma bacteriano tiene aproximadamente la misma probabilidad de ser transferido desde una bacteria dadora a una recipiente. La transduccin especializada difiere de la transduccin generalizada en varios puntos. Es mediada slo por fagos temperados especficos, y slo unos pocos genes dadores especficos pueden ser transferidos a la bacteria recipiente. Los fagos transductantes especializados son formados slo cuando la bacteria lisognica dadora entra el ciclo ltico, liberando la progenie de fagos.
Conjugacin: En la conjugacin, el contacto celular entre la bacteria dadora y recipiente conduce al establecimiento de un puente citoplasmtico entre ambas y transfiere parte o todo el genoma de la bacteria dadora a la recipiente. La habilidad de la bacteria dadora est determinada por los plsmidos conjugativos, llamados tambin plsmidos de fertilidad o plsmidos sexuales. El plsmido F (tambin factor F) de E. coli, es el prototipo para los plsmido de fertilidad en bacterias Gram (-). Las cepas de E. coli con un plsmido F extracromosomal es llamado F+ y funciona como dadores de ADN. En cambio, las cepas que carecen del plsmido F son denominados F- y se comportan como recipientes. Las funciones conjugativas de los plsmidos D son especificados por un cluster de al menos 25 genes de transferencia (tra) los cuales determinan la expresin del pili sexual, sntesis y transferencia del ADN durante el apareamiento, interferencia con la habilidad de las bacterias F+ para servir como recipiente, y otras funciones. Cada bacteria F+ tiene de 1 a 3 pili sexuales que se unen a una protena de membrana externa especfica en la bacteria recipiente. Luego se forma un puente citoplasmtico intercelular y una hebra del ADN del plsmido F es transferido desde la bacteria dadora a la bacteria recipiente, comenzando en un sitio nico y progresando en la direccin 5 a 3. La hebra transferida a la bacteria recipiente es convertida a una doble hebra circular, constituyendo un nuevo plsmido F. El plsmido F en E. coli puede existir como un elemento extracromosmico o ser integrado en el cromosoide bacteriano. E. coli, posee mltiples copias de diferentes elementos genticos llamados secuencias de insercin (IS), en varios sitios de su cromosoide. Se ha observado que el plsmido F se integra preferentemente en aquellas regiones que contienen secuencias de insercin. Una cepa de E. coli con un plsmido F integrado mantiene su habilidad para funcionar como una dador de ADN. Debido a que las cepas con factores F integrado pueden transferir genes cromosomales a bacteria recipientes con una alta frecuencia, son llamadas cepas Hfr. La transferencia mediada por el factor F integrado, tambin involucra una sola hebra, pero en este caso el replicn es hbrido, pues se transfiere parte del ADN del factor F y parte del ADN del cromosoide bacteriano. Los plsmidos F integrados en cepas Hfr pueden algunas veces ser excendidos desde el cromosoide bacteriano. Si el proceso de excisin es preciso, se generan clulas F+ nuevamente. Sin embargo, en raras ocasiones la excisin no es perfecta y fragmentos del ADN bacteriano pueden ser incorporadas al plsmido F, formndose un replicn hbrido, llamado plsmido F.
38 Biologa de los microorganismos
La conjugacin tambin ocurre en bacterias Gram (+). La bacteria Gram (+), dadora de ADN, produce adhesinas que causan su agregacin con clulas recipientes, pero el pili sexual no est involucrado. En alguna especies de Streptococcus, la bacteria recipiente produce ferohormonas sexuales, extracelulares, que provocan que el fenotipo donador sea expresado en aquellas bacterias que contienen un plsmido conjugativo apropiado.
BIBLIOGRAFA
Griffiths, A., Miller, J., Suzuki, D., Lewontin, R. and Gelbart, W. 1996. An Introduction to Genetic Analysis. Sixth Edition. W. H. Freeman and Company. New York. U.S.A. Murray, P, Kobayashi, G. Pfaller, M. and Rosenthal, K. 1997. Microbiologa Mdica. Segunda edicin. Harcourt Brace de Espaa, S.A. Madrid, Espaa. Snyder, L. and Champness, W. 1997. Molecular Genetics of bacteria. American Society for Microbiology Press. Washington, U.S.A.
You might also like
- Transformacion de Tejidos de Papa Con Agrobacterium Tumefasciens Portadores Del Gen Cry 1aDocument2 pagesTransformacion de Tejidos de Papa Con Agrobacterium Tumefasciens Portadores Del Gen Cry 1aGiovani AltezNo ratings yet
- Genética I (T1 - T13)Document149 pagesGenética I (T1 - T13)Rocío Prieto VázquezNo ratings yet
- Transformación en E.coli, Extracción, Purificación y Digestión Del Plásmido pQE-30 + sGFP4 Con Un Posterior Análisis Mediante ElectroforesisDocument10 pagesTransformación en E.coli, Extracción, Purificación y Digestión Del Plásmido pQE-30 + sGFP4 Con Un Posterior Análisis Mediante ElectroforesisVíctor Segarra LindaoNo ratings yet
- EficienciaDocument2 pagesEficienciaFlavia PrietoNo ratings yet
- Transformación BacterianaDocument2 pagesTransformación BacterianajmoreiratorresNo ratings yet
- Transgénesis plantasDocument16 pagesTransgénesis plantasArturo AquinoNo ratings yet
- Preguntas Examen SeptiembreDocument31 pagesPreguntas Examen SeptiembreRocio Valverde PastorNo ratings yet
- UDI 7. Clonación de Ácidos Nucleicos.Document9 pagesUDI 7. Clonación de Ácidos Nucleicos.juan manuel molina ramosNo ratings yet
- Capitulo 13 Biotecnologia Audesirk La Vida en La TierraDocument22 pagesCapitulo 13 Biotecnologia Audesirk La Vida en La TierraMiisii Arzuaga0% (1)
- Transgenesis Vegetal Expo PDFDocument14 pagesTransgenesis Vegetal Expo PDFJossy Morales100% (1)
- BiotecnologiaDocument46 pagesBiotecnologiaSHEYLA ANHELA VILCA HUANCANo ratings yet
- Ecología HumanaDocument14 pagesEcología HumanaNancy FaranaNo ratings yet
- Biologia, Inmunología IIDocument32 pagesBiologia, Inmunología IILinda0% (1)
- Protocolos extracción ADN bacteriasDocument3 pagesProtocolos extracción ADN bacteriasJavier RodriguezNo ratings yet
- Microbiologia Semana 1-Semana 7Document34 pagesMicrobiologia Semana 1-Semana 7Katherine RibbeckNo ratings yet
- DNA Como Portador de La Información Genética.Document7 pagesDNA Como Portador de La Información Genética.mferraNo ratings yet
- Diagnostico Molecular: Adn Y ArnDocument50 pagesDiagnostico Molecular: Adn Y ArnSalomé MontenegroNo ratings yet
- InteriorDocument96 pagesInteriorKutufaNo ratings yet
- Biodegradación de Petróleo Por BacteriasDocument9 pagesBiodegradación de Petróleo Por BacteriasCharles GarPerNo ratings yet
- P010 PlasmidorecombinanteDocument22 pagesP010 PlasmidorecombinanteMARITZA PALOMINO CAHUANANo ratings yet
- Genetica Bacteriana. Tema 16 VERSIÓN LARGADocument68 pagesGenetica Bacteriana. Tema 16 VERSIÓN LARGAMagdalena Oñate SerranoNo ratings yet
- Guía de Actividades y Rúbrica de Evaluación - Tarea 1 - Microorganismos ProcariotasDocument10 pagesGuía de Actividades y Rúbrica de Evaluación - Tarea 1 - Microorganismos ProcariotasERiKA REYESNo ratings yet
- Documento 14Document7 pagesDocumento 14Chabahi SidlbachirNo ratings yet
- Aplicaciones de La Ingeniería Genética y La Genómica en EnologíaDocument9 pagesAplicaciones de La Ingeniería Genética y La Genómica en EnologíaanaiNo ratings yet
- V Trabajo de Investigación de Biología Flor Villasmil 5to Año Sección C (3er Lapso)Document16 pagesV Trabajo de Investigación de Biología Flor Villasmil 5to Año Sección C (3er Lapso)Arianna CardenasNo ratings yet
- Capitulo 7 Segunda ParteDocument8 pagesCapitulo 7 Segunda ParteArelys GonzalónNo ratings yet
- PRACTICAS GeneticaDocument35 pagesPRACTICAS GeneticaMaria Alejandra SantanderNo ratings yet
- Cultivo de Tejidos VegetalesDocument15 pagesCultivo de Tejidos VegetalesAntonio TorresNo ratings yet
- Ingenieria GeneticaDocument24 pagesIngenieria Geneticaicrg0% (1)
- Generalidades Genetica MicrobianaDocument15 pagesGeneralidades Genetica MicrobianaCaroline Toledo PazmiñoNo ratings yet