You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- What Are The Importance of Genetics in AgricultureDocument7 pagesWhat Are The Importance of Genetics in AgricultureMaryam AliNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- S-B-5-3 - From Gene To Protein KEYDocument5 pagesS-B-5-3 - From Gene To Protein KEYfirebot4100% (1)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- C Value ParadoxDocument2 pagesC Value ParadoxRohit MaliNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Mto 201615Document7 pagesMto 201615Tim TiemannNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Chapter V: LIPIDSDocument23 pagesChapter V: LIPIDSGwyneth Marie DayaganNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Genomic and Non-Genomic Effects of Androgens in The Cardiovascular System: Clinical ImplicationsDocument14 pagesGenomic and Non-Genomic Effects of Androgens in The Cardiovascular System: Clinical ImplicationsGeraldine Villamil JiménezNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Multiple Choice On Proteins - MCQ Biology - Learning Biology Through MCQsDocument5 pagesMultiple Choice On Proteins - MCQ Biology - Learning Biology Through MCQsCésar Antonio Antonio RodriguezNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Lesson 5 - Central Dogma of Molecular BiologyDocument33 pagesLesson 5 - Central Dogma of Molecular BiologySherwin YenogacioNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Enzymes Amylase, Proteases, Lipases, and Cellulase: Applications and ManufacturersDocument3 pagesEnzymes Amylase, Proteases, Lipases, and Cellulase: Applications and ManufacturersKarl Anne DomingoNo ratings yet
- Catabolite RepressionDocument7 pagesCatabolite RepressionLucy SirinehNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- 1.1 Enzymology (Bravo)Document11 pages1.1 Enzymology (Bravo)Arman Carl DulayNo ratings yet
- Polymers: Acids PhosphateDocument10 pagesPolymers: Acids PhosphategqsdjztkydNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Biological Importance of MagnasiumDocument27 pagesBiological Importance of Magnasiumwarrierabhay100% (1)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- 10 1134@S102279541907010XDocument21 pages10 1134@S102279541907010X0xcffaaNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
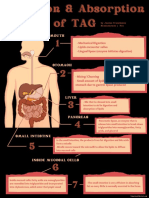
- Digestion & Absorption of TAGDocument1 pageDigestion & Absorption of TAGJanine Franchesca SuministradoNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Biological Revolution: Periodicals of Engineering and Natural Sciences Vol. 3 No. 1 (2015) Available Online atDocument2 pagesThe Biological Revolution: Periodicals of Engineering and Natural Sciences Vol. 3 No. 1 (2015) Available Online atAngel TaylorNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Reverse PharmacognosyDocument15 pagesReverse PharmacognosyJesús GutiérrezNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- Lab #06Document22 pagesLab #06ldlewisNo ratings yet
- Protein: Lecture #10 in Clinical Chemistry Lecture By: Hildegard Ariadne T. Coloma, RMT, MSMTDocument34 pagesProtein: Lecture #10 in Clinical Chemistry Lecture By: Hildegard Ariadne T. Coloma, RMT, MSMTJesley Buenaventura100% (1)
- QPCR vs. Digital PCR vs. Traditional PCRDocument4 pagesQPCR vs. Digital PCR vs. Traditional PCRnavnaNo ratings yet
- Biology Investigatory ProjectDocument12 pagesBiology Investigatory ProjectArnab86% (7)
- Product Support Booklet: Lara Briden Endometriosis and PcosDocument24 pagesProduct Support Booklet: Lara Briden Endometriosis and PcosVeronica TestaNo ratings yet
- Transfusion 2015 ChapuyDocument11 pagesTransfusion 2015 ChapuyNedelcu CarmenNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Enzymes PresDocument34 pagesEnzymes Presm. uma deviNo ratings yet
- 2015 Revision Tutorial Core Topic 2 and 4 AnsDocument35 pages2015 Revision Tutorial Core Topic 2 and 4 AnsliveaqNo ratings yet
- Explore The Mechanism of Liyin Decoction in Treating Nonalcoholic Steatohepatitis and Atherosclerosis With Homotherapy For Heteropathy Based On Network Pharmacology and Molecular DockingDocument13 pagesExplore The Mechanism of Liyin Decoction in Treating Nonalcoholic Steatohepatitis and Atherosclerosis With Homotherapy For Heteropathy Based On Network Pharmacology and Molecular DockingHerald Scholarly Open AccessNo ratings yet
- BIOL200 PSet - Unit 2Document17 pagesBIOL200 PSet - Unit 2Lovey ChandiNo ratings yet
- Energy For All!: Engage: Food For CornDocument4 pagesEnergy For All!: Engage: Food For CornBri LavaeNo ratings yet
- Muhammad Wajid: Institute of Pharmacy Gulab Devi Educational Complex, LahoreDocument31 pagesMuhammad Wajid: Institute of Pharmacy Gulab Devi Educational Complex, LahoreAhmed ImranNo ratings yet
- Calder 2013Document34 pagesCalder 2013Lastrie BuluatieNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)