You might also like
- Banco de Preguntas de Biologia MolecularDocument31 pagesBanco de Preguntas de Biologia MolecularFrancisco Rodriguez100% (1)
- Diseño Molecular de La Vida PDFDocument6 pagesDiseño Molecular de La Vida PDFTania JacintoNo ratings yet
- PRESENT SIMPLE EN INGLÉSDocument3 pagesPRESENT SIMPLE EN INGLÉSAndres Pardo100% (1)
- Cuestionario AcDocument9 pagesCuestionario AcVirginia De La CruzNo ratings yet
- Metabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesFrom EverandMetabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesNo ratings yet
- Nucleósidos y NucleótidosDocument2 pagesNucleósidos y NucleótidosAdriana RodriguezNo ratings yet
- Proteínas: estructura, clasificación, funciones y fuentes alimentariasDocument5 pagesProteínas: estructura, clasificación, funciones y fuentes alimentariasSteeven Espinosa Otavalo100% (1)
- Examen bioquímica falso verdaderoDocument2 pagesExamen bioquímica falso verdaderoYuMi BautistaNo ratings yet
- Paper EnzimasDocument5 pagesPaper EnzimasErick VillaNo ratings yet
- Bioquimica Del Agua y Tampones FisiologicosDocument24 pagesBioquimica Del Agua y Tampones FisiologicospedrojacintohNo ratings yet
- Cuestionario AminoacidosDocument8 pagesCuestionario AminoacidosMichael Hernandez33% (3)
- PROTEÍNASDocument14 pagesPROTEÍNASJose Maravi100% (2)
- Agua BioquímicaDocument99 pagesAgua BioquímicalupitagroNo ratings yet
- Biomoléculas - ProteínasDocument24 pagesBiomoléculas - ProteínasAdriana EliNo ratings yet
- Caracterìsticas y Funciones de Las Células Del SIDocument107 pagesCaracterìsticas y Funciones de Las Células Del SISara Elizabeth CisnerosNo ratings yet
- PROTEÍNASDocument39 pagesPROTEÍNASkatia100% (1)
- CelulaDocument27 pagesCelulaFransis Gonçalves100% (2)
- Fosforilacion OxidativaDocument27 pagesFosforilacion Oxidativajosuekstillo100% (4)
- Metabolismo proteínas aminoácidos UNCPDocument21 pagesMetabolismo proteínas aminoácidos UNCPRonald Astucuri PalaciosNo ratings yet
- Metabolismo de Nucleotidos.10Document24 pagesMetabolismo de Nucleotidos.10Edwar Tarrillo Vasquez100% (1)
- Bioquimica de ProteinasDocument160 pagesBioquimica de ProteinasChristian CastilloNo ratings yet
- Las ProteínasDocument4 pagesLas ProteínasJuan Anillares100% (1)
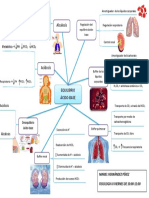
- Regulación del equilibrio ácido-base a través de los sistemas respiratorio y renalDocument1 pageRegulación del equilibrio ácido-base a través de los sistemas respiratorio y renalMariel Hernândez Pêrez100% (1)
- ALQUINOSDocument12 pagesALQUINOSsilviaNo ratings yet
- Fisiopatología del ERGE: Relajaciones del EEI causan reflujoDocument26 pagesFisiopatología del ERGE: Relajaciones del EEI causan reflujoJuan Diego Vilca PinedaNo ratings yet
- Aminoacidos y ProteinasDocument52 pagesAminoacidos y ProteinasElver Osorio100% (1)
- BiomoléculasArias PDFDocument584 pagesBiomoléculasArias PDFAndrea Morales100% (1)
- Esquema de Agua en El OrganismoDocument2 pagesEsquema de Agua en El OrganismoJovanna RivasNo ratings yet
- Fosforilación OxidativaDocument25 pagesFosforilación OxidativaEmilio Jeanpaul Mitacc GutierrezNo ratings yet
- Disacáridos: Sacarosa, Lactosa y MaltosaDocument35 pagesDisacáridos: Sacarosa, Lactosa y MaltosaLuis Alberto Flores ParedesNo ratings yet
- Equilibrio HidroelectroliticoDocument21 pagesEquilibrio HidroelectroliticomiguelNo ratings yet
- Metabolismo de Las ProteinasDocument18 pagesMetabolismo de Las ProteinasLuis CamaraNo ratings yet
- Clase 1. El AguaDocument47 pagesClase 1. El AguaNilts Vega CuyaNo ratings yet
- Grasas y LípidosDocument26 pagesGrasas y LípidosAlonsoNo ratings yet
- Cadena RespiratoriaDocument2 pagesCadena RespiratoriaJesus RamirezNo ratings yet
- Beta OxidacionDocument20 pagesBeta OxidacionJâzmïn ZårcöNo ratings yet
- OperonDocument2 pagesOperonDaniiel VillegasNo ratings yet
- CarbohidratosDocument28 pagesCarbohidratosAnonimo22100% (2)
- Cilios EstereociliosDocument3 pagesCilios EstereociliosLeón Felipe López MartínezNo ratings yet
- Reporte y Series de Casos - ArtículoDocument6 pagesReporte y Series de Casos - Artículoerika beltranNo ratings yet
- LÍPIDOSDocument10 pagesLÍPIDOSRafael Angel Huiza Trujillo100% (3)
- Digestión y Absorción de ProteínasDocument5 pagesDigestión y Absorción de ProteínasclaudiaNo ratings yet
- PROTEINASDocument26 pagesPROTEINASMariselCielo100% (1)
- Fosforilacion OxidativaDocument11 pagesFosforilacion OxidativaJose Alberto Ariza PasoNo ratings yet
- 1-07 Acidos NucléicosDocument55 pages1-07 Acidos NucléicosDan EmpleoNo ratings yet
- Digestión y absorción de lípidosDocument16 pagesDigestión y absorción de lípidosEros RamazottiNo ratings yet
- Resúmen Fisiología EndócrinoDocument16 pagesResúmen Fisiología EndócrinoWalter Wellmann CatalánNo ratings yet
- Bases Moleculares de La Información GenéticaDocument13 pagesBases Moleculares de La Información GenéticaIrene Sánchez LouNo ratings yet
- Bomba Sodio PotasioDocument31 pagesBomba Sodio PotasioAndres Felipe Celis CallejasNo ratings yet
- NUCLEÓTIDOSDocument5 pagesNUCLEÓTIDOSGaby_biologia83% (6)
- Errores metabólicos homocistinuriaDocument45 pagesErrores metabólicos homocistinuriaMari CarmenNo ratings yet
- Metabolismo celular: glucólisis y ciclo de KrebsDocument61 pagesMetabolismo celular: glucólisis y ciclo de KrebsAndres Monroy100% (1)
- Difusion y OsmosisDocument8 pagesDifusion y OsmosisAndres BolivarNo ratings yet
- Eritropoyetina, regulador de la eritropoyesisDocument2 pagesEritropoyetina, regulador de la eritropoyesisLupiita RamirezNo ratings yet
- ENZIMASDocument59 pagesENZIMASÉrÌck PÂrédèsNo ratings yet
- ELECTROLITOSDocument62 pagesELECTROLITOSYuleen ZambranoNo ratings yet
- Citoquinas: Proteínas clave del sistema inmuneDocument3 pagesCitoquinas: Proteínas clave del sistema inmuneAbel CamachoNo ratings yet
- El CitoplasmaDocument2 pagesEl CitoplasmaDavid Gato100% (1)
- AlbúminaDocument5 pagesAlbúminaGabriel WardNo ratings yet
- Indicadores BioquímicosDocument151 pagesIndicadores BioquímicosRodrigo Romero PichardoNo ratings yet
- Presentacion Fisiologia HemoglobinaDocument6 pagesPresentacion Fisiologia HemoglobinaManuel SepulvedaNo ratings yet
- MoralDocument8 pagesMoralKas AndradeNo ratings yet
- Quimica NivelaciónDocument2 pagesQuimica NivelaciónJordan AguilarNo ratings yet
- Quimica GasesDocument2 pagesQuimica GasesJordan AguilarNo ratings yet
- Quimica NivelaciónDocument2 pagesQuimica NivelaciónJordan AguilarNo ratings yet
- Quimica GasesDocument2 pagesQuimica GasesJordan AguilarNo ratings yet
- Trabajo de CalculoDocument4 pagesTrabajo de CalculoJordan AguilarNo ratings yet
- Jordan TecnologiaDocument9 pagesJordan TecnologiaJordan AguilarNo ratings yet
- El Comport A Mien To de Los GasesDocument6 pagesEl Comport A Mien To de Los GasesJordan AguilarNo ratings yet
- Terminos TecnologiaDocument27 pagesTerminos TecnologiaJordan AguilarNo ratings yet
- EnvironmentDocument31 pagesEnvironmentJordan AguilarNo ratings yet
- Terminos Tecnologia #IDocument2 pagesTerminos Tecnologia #IJordan AguilarNo ratings yet
- Duplicacion de AdnDocument4 pagesDuplicacion de AdnJordan AguilarNo ratings yet
- Replicacion de AdnDocument3 pagesReplicacion de AdnJordan AguilarNo ratings yet
- Sintesis de ADNDocument20 pagesSintesis de ADNJordan Aguilar88% (8)
- Replicacion de AdnDocument3 pagesReplicacion de AdnJordan AguilarNo ratings yet
- FuturismoDocument7 pagesFuturismoJordan AguilarNo ratings yet
- AlcoholismDocument12 pagesAlcoholismJordan AguilarNo ratings yet
- Bmo 46 LegastDocument19 pagesBmo 46 LegastJuan Carlos TrianaNo ratings yet
- Cientifics ArticlesDocument6 pagesCientifics ArticlesJordan AguilarNo ratings yet
- Formato Matricula 2011 2Document1 pageFormato Matricula 2011 2Jordan AguilarNo ratings yet
- Calendario Academico II 2011Document1 pageCalendario Academico II 2011Jordan AguilarNo ratings yet
- Curriculum VitaeDocument2 pagesCurriculum VitaeJordan AguilarNo ratings yet
- El YageDocument15 pagesEl YageJordan AguilarNo ratings yet
- Horarios II 11Document12 pagesHorarios II 11Jordan AguilarNo ratings yet
- CochasDocument42 pagesCochasJordan AguilarNo ratings yet
- Abstract ArtDocument14 pagesAbstract ArtJordan AguilarNo ratings yet
- Las Preguntas Del MaltratoDocument2 pagesLas Preguntas Del MaltratoJordan AguilarNo ratings yet
- Look Forward ToDocument6 pagesLook Forward ToJordan AguilarNo ratings yet
- Clonación Del Gen Del Factor VIII de Coagulación HumanoDocument5 pagesClonación Del Gen Del Factor VIII de Coagulación HumanomczanaboniNo ratings yet
- Rep 312 PDFDocument253 pagesRep 312 PDFchaquila22No ratings yet
- Unidad 16 - Transcripcion y TraducciónDocument19 pagesUnidad 16 - Transcripcion y TraducciónLaura MoyaNo ratings yet
- Carbohidratos 2Document22 pagesCarbohidratos 2Myriam Castillo AlonzoNo ratings yet
- Taller de Recuperacion Biologia Celular y MolecularDocument7 pagesTaller de Recuperacion Biologia Celular y MolecularJuanEstebanMoraNo ratings yet
- La Replicación de Los PrionesDocument2 pagesLa Replicación de Los PrionesshiNo ratings yet
- Articulo Martinez Miguel JenniferDocument47 pagesArticulo Martinez Miguel Jenniferkatherin PCNo ratings yet
- Genética Del Desarrollo EmbrionarioDocument3 pagesGenética Del Desarrollo EmbrionarioLector ÁvidoNo ratings yet
- Documento PDFDocument1 pageDocumento PDFmargarita oshea gumucioNo ratings yet
- Tarea .: Nombre: Cáceres Mesías Nohely Nivel 1 Selección 3 (2)Document7 pagesTarea .: Nombre: Cáceres Mesías Nohely Nivel 1 Selección 3 (2)Nohely Cáceres MesíasNo ratings yet
- Estructura GFP 38Document2 pagesEstructura GFP 38Melkid Yums Esquen BayonaNo ratings yet
- 09-08-2021 Enzimas Reguladoras-Ing. QuimicaDocument30 pages09-08-2021 Enzimas Reguladoras-Ing. QuimicaLeoncio Hilario UmiyauriNo ratings yet
- ENZIMASDocument27 pagesENZIMASEdwin J MorenoNo ratings yet
- SP 10Document3 pagesSP 10Sandra Nicol MarquinaorbegozoNo ratings yet
- EBAU Biología Bloque 1: Componentes y estructura de la vidaDocument2 pagesEBAU Biología Bloque 1: Componentes y estructura de la vidaRayz 2004 xNo ratings yet
- Replicacion Del ADNDocument5 pagesReplicacion Del ADNMARY RodríguezNo ratings yet
- Informe de LipoproteínasDocument16 pagesInforme de LipoproteínasFiorelaMHNo ratings yet
- Presentacion Arn y AdnDocument9 pagesPresentacion Arn y AdnalidamontenegrodiazNo ratings yet
- Cinética enzimática: modelos y ecuacionesDocument11 pagesCinética enzimática: modelos y ecuacionesDiana Alexandra Santos RiveraNo ratings yet
- Las Proteinas ExposicionDocument8 pagesLas Proteinas ExposicionKatherin Pulido100% (1)
- Tirosinasa 2Document5 pagesTirosinasa 2Daniela Valencia LópezNo ratings yet
- PCR III AKBioq-2017-parte2Document18 pagesPCR III AKBioq-2017-parte2Therry JordanNo ratings yet
- Dogma de La Biología Celular. Estructura de Un Gen. Regulación de La Expresión Génica y Código GenéticoDocument16 pagesDogma de La Biología Celular. Estructura de Un Gen. Regulación de La Expresión Génica y Código GenéticoDaniel RojasNo ratings yet
- Niveles de Organización de Las ProteínasDocument2 pagesNiveles de Organización de Las ProteínasMero1053No ratings yet
- La estructura del ADN y su función en la herencia genéticaDocument18 pagesLa estructura del ADN y su función en la herencia genéticabrian alburquequeNo ratings yet
- ILLUMINA ResumenDocument4 pagesILLUMINA ResumenJenny GonzalezNo ratings yet
- Bacterio Fag OsDocument3 pagesBacterio Fag OsSandra QuintanaNo ratings yet
- Minuta Sobre ProteinasDocument10 pagesMinuta Sobre Proteinasdaneicy volcanesNo ratings yet