You might also like
- Probiotic Rescue: How You can use Probiotics to Fight Cholesterol, Cancer, Superbugs, Digestive Complaints and MoreFrom EverandProbiotic Rescue: How You can use Probiotics to Fight Cholesterol, Cancer, Superbugs, Digestive Complaints and MoreNo ratings yet
- Microbes as Feed Additives for LivestockDocument7 pagesMicrobes as Feed Additives for LivestockMd Manirul IslamNo ratings yet
- Fermented Foods for Health: Use the Power of Probiotic Foods to Improve Your Digestion, Strengthen Your Immunity, and Prevent IllnessFrom EverandFermented Foods for Health: Use the Power of Probiotic Foods to Improve Your Digestion, Strengthen Your Immunity, and Prevent IllnessRating: 4.5 out of 5 stars4.5/5 (4)
- 4w100002599 PDFDocument8 pages4w100002599 PDFPrakashNo ratings yet
- Fabulous Fermentation: A Comprehensive Guide to The Health Benefits of FermentationFrom EverandFabulous Fermentation: A Comprehensive Guide to The Health Benefits of FermentationNo ratings yet
- Effect of Probiotics/Prebiotics On Cattle Health and ProductivityDocument7 pagesEffect of Probiotics/Prebiotics On Cattle Health and ProductivityJuan Manuel Cabrales FloresNo ratings yet
- Gut Check: Unleash the Power of Your Microbiome to Reverse Disease and Transform Your Mental, Physical, and Emotional Health...SummarizedFrom EverandGut Check: Unleash the Power of Your Microbiome to Reverse Disease and Transform Your Mental, Physical, and Emotional Health...SummarizedNo ratings yet
- Beneficial Bacteria in Pet Rabbits: DigestionDocument8 pagesBeneficial Bacteria in Pet Rabbits: DigestionHouse HuntingNo ratings yet
- Yogurt as a Probiotic Carrier FoodDocument17 pagesYogurt as a Probiotic Carrier Foodsilver_10882698No ratings yet
- Microflora of Digestive Tract in PoultryDocument4 pagesMicroflora of Digestive Tract in Poultrymaria fernandaNo ratings yet
- Probiotics From Human Breast Milk and Their ApplicationDocument22 pagesProbiotics From Human Breast Milk and Their ApplicationStephen MooreNo ratings yet
- Human Probiotics and PrebioticsDocument32 pagesHuman Probiotics and PrebioticsVijayaraghavan GonuguntlaNo ratings yet
- Probio PCCDocument8 pagesProbio PCCCherry San DiegoNo ratings yet
- Animals: Gut Microbiota-Polyphenol Interactions in Chicken: A ReviewDocument18 pagesAnimals: Gut Microbiota-Polyphenol Interactions in Chicken: A ReviewevelNo ratings yet
- ProbioticsDocument35 pagesProbioticsJavier HernandezNo ratings yet
- Effects of Prebiotics, Probiotics and Synbiotics in The Diet of Young PigsDocument179 pagesEffects of Prebiotics, Probiotics and Synbiotics in The Diet of Young Pigspedro41100% (1)
- Shigella FlexneryDocument29 pagesShigella FlexneryFernandaAbahonzaNo ratings yet
- 1 Intestinal Fermentation: Dietary and Microbial InteractionsDocument29 pages1 Intestinal Fermentation: Dietary and Microbial InteractionsFernandaAbahonzaNo ratings yet
- BiavatiB Probiotics and PrebioticsDocument75 pagesBiavatiB Probiotics and PrebioticsRashmin ShahNo ratings yet
- The Impact of Probiotic On Gut Health: Current Drug Metabolism February 2009Document12 pagesThe Impact of Probiotic On Gut Health: Current Drug Metabolism February 2009simalihaNo ratings yet
- Potential of Probiotic Lactobacillus Strains As Food AdditivesDocument16 pagesPotential of Probiotic Lactobacillus Strains As Food AdditivesSheetalPithvaNo ratings yet
- Aj 1Document11 pagesAj 1Trần Tấn TàiNo ratings yet
- Abiotic, Biotic and ProbioticsDocument4 pagesAbiotic, Biotic and ProbioticsSiddNo ratings yet
- prebioticsDocument6 pagesprebioticsBarkat IbtissamNo ratings yet
- Intestinal Microbiota in Human Health and Disease: The Impact of ProbioticsDocument32 pagesIntestinal Microbiota in Human Health and Disease: The Impact of ProbioticsSharan SahotaNo ratings yet
- Lecture 4 - ProbioticsDocument38 pagesLecture 4 - Probioticsda6220848No ratings yet
- World Gastroenterology Organisation Global Guidelines - SafetyDocument28 pagesWorld Gastroenterology Organisation Global Guidelines - SafetyDianaNo ratings yet
- 1989 - Fuller R. - Probiotics in Man and AnimalsDocument14 pages1989 - Fuller R. - Probiotics in Man and AnimalsFelipe ZapataNo ratings yet
- Lactobalciusn DentalDocument10 pagesLactobalciusn Dentallia apriloaNo ratings yet
- Human Fecal Flora11Document15 pagesHuman Fecal Flora11hussainNo ratings yet
- A-The Importance of Probiotic Institut Rosell Inc. (Montreal, QCDocument4 pagesA-The Importance of Probiotic Institut Rosell Inc. (Montreal, QCAMIT JAINNo ratings yet
- Probiotics Review and Future AspectsDocument5 pagesProbiotics Review and Future AspectsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- 1 s2.0 S1756464615002121 MainDocument12 pages1 s2.0 S1756464615002121 MainDuzzysNo ratings yet
- Ensayos: Criterios de Calidad de Los Microorganismos Probióticos Y Evidencias Sobre Efectos HipocolesterolémicosDocument8 pagesEnsayos: Criterios de Calidad de Los Microorganismos Probióticos Y Evidencias Sobre Efectos HipocolesterolémicosJavier Lebron MoralesNo ratings yet
- Probiotics and PrebioticsDocument6 pagesProbiotics and Prebioticsrangga hilmiNo ratings yet
- The Benefits of Gene TherapyDocument3 pagesThe Benefits of Gene TherapyLevi AckermanNo ratings yet
- Microbiota PerrosDocument12 pagesMicrobiota PerrosJesus BurtonNo ratings yet
- 59 170 1 PBDocument4 pages59 170 1 PBBaja HitamNo ratings yet
- Gastrointestinal Microflora PPT 19.1.11.Document51 pagesGastrointestinal Microflora PPT 19.1.11.सुमेर सिंह मीणाNo ratings yet
- Introduction of Colonic and Fecal Microbiota FromDocument16 pagesIntroduction of Colonic and Fecal Microbiota FromEduardo ViolaNo ratings yet
- Fish & Shell Fish Immunology: S.K. NayakDocument13 pagesFish & Shell Fish Immunology: S.K. NayakYasir MahmoodNo ratings yet
- Makalah 2Document8 pagesMakalah 2ainanNo ratings yet
- Prebiotics in Companion and Livestock AnDocument112 pagesPrebiotics in Companion and Livestock AnIonela HoteaNo ratings yet
- Review: Roles of Prebiotics in Intestinal Ecosystem of BroilersDocument18 pagesReview: Roles of Prebiotics in Intestinal Ecosystem of Broilersamamùra maamarNo ratings yet
- Diet Composition and Non-Antibiotic Feeding Programs in EuropeDocument14 pagesDiet Composition and Non-Antibiotic Feeding Programs in EuropeDalia MatjodNo ratings yet
- Streptococcus, and Bacillus. Sequence Alignment of These Putative Bacteriocins Revealed That TheDocument2 pagesStreptococcus, and Bacillus. Sequence Alignment of These Putative Bacteriocins Revealed That ThemaxNo ratings yet
- Effects of Diets Containing Different ConcentratioDocument11 pagesEffects of Diets Containing Different ConcentratioSuci EkoNo ratings yet
- Fal Cinelli 2018Document7 pagesFal Cinelli 2018SEBASTIAN JOSE VEGA MARCELONo ratings yet
- BAMN11 ProbioticsDocument4 pagesBAMN11 ProbioticsSUBRATA PARAINo ratings yet
- Probiotics Research PaperDocument12 pagesProbiotics Research Paperapi-271090706No ratings yet
- Direct Fed MicrobialsDocument4 pagesDirect Fed MicrobialsSubrata ParaiNo ratings yet
- Kabluchko - 2017 - SURVIVAL OF MICROORGANISMS FROM MODERN PROBIOTICS IN MODEL CONDITIONS OF THE INTESTINEDocument6 pagesKabluchko - 2017 - SURVIVAL OF MICROORGANISMS FROM MODERN PROBIOTICS IN MODEL CONDITIONS OF THE INTESTINEMamen Casado MuñozNo ratings yet
- Food Research InternationalDocument10 pagesFood Research InternationalSandraJSantosNo ratings yet
- 10 Iajps10062017 PDFDocument10 pages10 Iajps10062017 PDFBaru Chandrasekhar RaoNo ratings yet
- Pro Bio Tics in Pediatric PracticeDocument48 pagesPro Bio Tics in Pediatric PracticeOLUKAYODEVNo ratings yet
- Impact of Banana Puree On Viability of P PDFDocument8 pagesImpact of Banana Puree On Viability of P PDFAndreea FlorentinaNo ratings yet
- Bacillus Subtilis Litopenaeus Vannamei: Effect of On The Growth and Survival Rate of ShrimpDocument8 pagesBacillus Subtilis Litopenaeus Vannamei: Effect of On The Growth and Survival Rate of ShrimpKalyanaraman MahadevanNo ratings yet
- Effect of Probiotic Supplementation on the Performance of BroilersDocument7 pagesEffect of Probiotic Supplementation on the Performance of BroilersPrem RajNo ratings yet
- Potential Use of Probiotics: Ekachai ChukeatiroteDocument8 pagesPotential Use of Probiotics: Ekachai ChukeatiroteDanu EffendiNo ratings yet
- Overview of Differences Between MicrobiaDocument10 pagesOverview of Differences Between MicrobiaAlanGonzalezNo ratings yet
- Digoxin FABDocument6 pagesDigoxin FABqwer22No ratings yet
- Cartha Worth SharingDocument27 pagesCartha Worth SharingtereAC85No ratings yet
- Group 1 RDL2Document101 pagesGroup 1 RDL2ChristelNo ratings yet
- PNP P.A.T.R.O.L. 2030 Score Card Dashboard FormulationDocument89 pagesPNP P.A.T.R.O.L. 2030 Score Card Dashboard FormulationMark Payumo83% (41)
- Research Paper Is at DominosDocument6 pagesResearch Paper Is at Dominosssharma83No ratings yet
- 2009 IBP ElectionsDocument77 pages2009 IBP ElectionsBaldovino VenturesNo ratings yet
- A Systematic Literature Review of Empirical Research On ChatGPT in EducationDocument23 pagesA Systematic Literature Review of Empirical Research On ChatGPT in Educationgraciduttra.profNo ratings yet
- Lesson 6. TNCTDocument32 pagesLesson 6. TNCTEsther EdaniolNo ratings yet
- Veerabhadra Swamy MantrasDocument6 pagesVeerabhadra Swamy Mantrasगणेश पराजुलीNo ratings yet
- Unit 11 LeadershipDocument4 pagesUnit 11 LeadershipMarijana DragašNo ratings yet
- Bunga Refira - 1830104008 - Allophonic RulesDocument6 pagesBunga Refira - 1830104008 - Allophonic RulesBunga RefiraNo ratings yet
- Edu 510 Final ProjectDocument13 pagesEdu 510 Final Projectapi-324235159No ratings yet
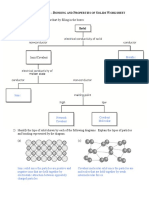
- Bonding and Properties of Solids Worksheet Solutions 1kadax6Document4 pagesBonding and Properties of Solids Worksheet Solutions 1kadax6Mel Patricia M. CabreraNo ratings yet
- Opinions and ThoughtsDocument2 pagesOpinions and Thoughtsfikri alfaroqNo ratings yet
- New Titles List 2014, Issue 1Document52 pagesNew Titles List 2014, Issue 1Worldwide Books CorporationNo ratings yet
- Hi Scan Pro ManualDocument231 pagesHi Scan Pro ManualFaridhul IkhsanNo ratings yet
- ML Performance Improvement CheatsheetDocument11 pagesML Performance Improvement Cheatsheetrahulsukhija100% (1)
- Metabical Positioning and CommunicationDocument15 pagesMetabical Positioning and CommunicationJSheikh100% (2)
- Module 3 in Oral Com 1Document20 pagesModule 3 in Oral Com 1Trisha DiohenNo ratings yet
- Jason A Brown: 1374 Cabin Creek Drive, Nicholson, GA 30565Document3 pagesJason A Brown: 1374 Cabin Creek Drive, Nicholson, GA 30565Jason BrownNo ratings yet
- Case Problems For Management ScienceDocument11 pagesCase Problems For Management ScienceMargarita AtanacioNo ratings yet
- Students Playwriting For Language DevelopmentDocument3 pagesStudents Playwriting For Language DevelopmentSchmetterling TraurigNo ratings yet
- Existentialism Is A HumanismDocument4 pagesExistentialism Is A HumanismAlex MendezNo ratings yet
- (123doc) - Internship-Report-Improving-Marketing-Strategies-At-Telecommunication-Service-Corporation-Company-VinaphoneDocument35 pages(123doc) - Internship-Report-Improving-Marketing-Strategies-At-Telecommunication-Service-Corporation-Company-VinaphoneK59 PHAN HA PHUONGNo ratings yet
- 04-DDD.Assignment 2 frontsheet 2018-2019-đã chuyển đổi PDFDocument21 pages04-DDD.Assignment 2 frontsheet 2018-2019-đã chuyển đổi PDFl1111c1anh-5No ratings yet
- Portal ScienceDocument5 pagesPortal ScienceiuhalsdjvauhNo ratings yet
- Richard Herrmann-Fractional Calculus - An Introduction For Physicists-World Scientific (2011)Document274 pagesRichard Herrmann-Fractional Calculus - An Introduction For Physicists-World Scientific (2011)Juan Manuel ContrerasNo ratings yet
- Ethical CRM PracticesDocument21 pagesEthical CRM Practicesanon_522592057No ratings yet
- Elderly Suicide FactsDocument2 pagesElderly Suicide FactsThe News-HeraldNo ratings yet
- Fish School 101: A Step-by-Step Guide for Fish OwnersFrom EverandFish School 101: A Step-by-Step Guide for Fish OwnersRating: 5 out of 5 stars5/5 (16)
- Livebearers: Understanding Guppies, Mollies, Swordtails and OthersFrom EverandLivebearers: Understanding Guppies, Mollies, Swordtails and OthersNo ratings yet
- Arowana: The Complete Owner’s Guide for the Most Expensive Fish in the World - Arowana Fish Tank, Types, Care, Food, Habitat, Breeding, Mythology – Includes Silver, Platinum, Red, Jardini, Black, Golden, GreenFrom EverandArowana: The Complete Owner’s Guide for the Most Expensive Fish in the World - Arowana Fish Tank, Types, Care, Food, Habitat, Breeding, Mythology – Includes Silver, Platinum, Red, Jardini, Black, Golden, GreenNo ratings yet
- Biofloc Fish Farming for Dummies: The Beginner's Guide to Setting Up FarmFrom EverandBiofloc Fish Farming for Dummies: The Beginner's Guide to Setting Up FarmNo ratings yet
- The Dragon Behind the Glass: A True Story of Power, Obsession, and the World's Most Coveted FishFrom EverandThe Dragon Behind the Glass: A True Story of Power, Obsession, and the World's Most Coveted FishRating: 4 out of 5 stars4/5 (35)
- Essential Tropical Fish Species Guide: The simplified guide to identifying, selecting and caring for your tropical fishFrom EverandEssential Tropical Fish Species Guide: The simplified guide to identifying, selecting and caring for your tropical fishNo ratings yet
- Sharks: 101 Super Fun Facts And Amazing Pictures (Featuring The World's Top 10 Sharks With Coloring Pages)From EverandSharks: 101 Super Fun Facts And Amazing Pictures (Featuring The World's Top 10 Sharks With Coloring Pages)Rating: 3 out of 5 stars3/5 (3)
- Dolphins: 101 Fun Facts & Amazing Pictures (Featuring The World's 6 Top Dolphins)From EverandDolphins: 101 Fun Facts & Amazing Pictures (Featuring The World's 6 Top Dolphins)Rating: 4 out of 5 stars4/5 (2)
- Understanding Life Below the Beautiful Sea | Children's Fish & Marine LifeFrom EverandUnderstanding Life Below the Beautiful Sea | Children's Fish & Marine LifeNo ratings yet
- The Big Book of Pet Names ~ More than 10,000 Pet Names! The Most Complete Guide to Pet Names & MeaningsFrom EverandThe Big Book of Pet Names ~ More than 10,000 Pet Names! The Most Complete Guide to Pet Names & MeaningsNo ratings yet
- Aquascaping: A Step-by-Step Guide to Planting, Styling, and Maintaining Beautiful AquariumsFrom EverandAquascaping: A Step-by-Step Guide to Planting, Styling, and Maintaining Beautiful AquariumsRating: 5 out of 5 stars5/5 (4)
- Avoid the 5 Most Common Saltwater Aquarium MistakesFrom EverandAvoid the 5 Most Common Saltwater Aquarium MistakesRating: 5 out of 5 stars5/5 (1)
- 5th Grade Geography: Seas and Oceans of the World: Fifth Grade Books Marine Life and Oceanography for KidsFrom Everand5th Grade Geography: Seas and Oceans of the World: Fifth Grade Books Marine Life and Oceanography for KidsNo ratings yet
- The New Saltwater Aquarium Guide: How to Care for and Keep Marine Fish and CoralsFrom EverandThe New Saltwater Aquarium Guide: How to Care for and Keep Marine Fish and CoralsRating: 5 out of 5 stars5/5 (2)