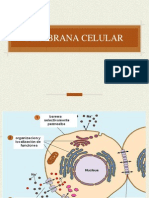
You might also like
- Semana 1 (Teoria)Document23 pagesSemana 1 (Teoria)Diego Alonso Segundo Diaz HinostrozaNo ratings yet
- Estructura de La MembranaDocument15 pagesEstructura de La MembranaSu SaucedoNo ratings yet
- MEMBRANA CELULAR Y TRANSPORTE Original CompletoDocument11 pagesMEMBRANA CELULAR Y TRANSPORTE Original CompletoFelicitasNo ratings yet
- Membrana Plasmatica MosaicoDocument3 pagesMembrana Plasmatica MosaicoAngie Melissa Franco RiveraNo ratings yet
- TAREA11Document9 pagesTAREA11Paula LopezNo ratings yet
- Cap 10 Membranas FinalDocument68 pagesCap 10 Membranas FinalPatricia Bustelo0% (1)
- Membranas celulares: estructura y funcionesDocument163 pagesMembranas celulares: estructura y funcionesStefani AtlleNo ratings yet
- Modulo 05 - Membranas BiologicasDocument17 pagesModulo 05 - Membranas BiologicasTamara Menéndez100% (1)
- Membrana Celular ProyectoDocument17 pagesMembrana Celular ProyectoGabriel Enrique Giler ReyesNo ratings yet
- 2.constituyentes de Las Membranas BiológicasDocument4 pages2.constituyentes de Las Membranas Biológicas90088264No ratings yet
- Guia 2 Biologia 10°°Document11 pagesGuia 2 Biologia 10°°Luciana pNo ratings yet
- Unidad 4Document25 pagesUnidad 4Jaqueline Alexias LozanoNo ratings yet
- Composición de Las Membranas BiológicasDocument7 pagesComposición de Las Membranas BiológicasAlberto SantanaNo ratings yet
- TEMA 8 Membrana Plasmatica y Organulos MembranososDocument22 pagesTEMA 8 Membrana Plasmatica y Organulos MembranososARIADNA MÉNDEZ ANCHUELONo ratings yet
- Resumen MembranaDocument15 pagesResumen MembranabelenNo ratings yet
- Las Membranas de La Célula Ejercen Diversas ActividadesDocument13 pagesLas Membranas de La Célula Ejercen Diversas ActividadesKauely SouzaNo ratings yet
- Guia de Estudios 5.y.6 .Leyva Vallejo Saul Arturo - Grupo 1Document14 pagesGuia de Estudios 5.y.6 .Leyva Vallejo Saul Arturo - Grupo 1arturoleyva2001No ratings yet
- Tema 7 Envolturas CelularesDocument16 pagesTema 7 Envolturas CelularesElena Rodriguez BejaranoNo ratings yet
- Modelo de Mosaico FluidoDocument7 pagesModelo de Mosaico FluidoIsabella Castro BerdugoNo ratings yet
- Membrana Plasmatica y CitoplasmicaDocument13 pagesMembrana Plasmatica y CitoplasmicaAlvaro Javier Perez PeñafielNo ratings yet
- Bioquimica Ii - Lipido-Qf-2023-IiDocument75 pagesBioquimica Ii - Lipido-Qf-2023-IiJoselin Milagros Ramos HuancaNo ratings yet
- 6 - BiomembranasDocument17 pages6 - BiomembranasFlor BovettiNo ratings yet
- Biología celular: Teoría celular y estructuraDocument13 pagesBiología celular: Teoría celular y estructuraCarmen Rosario Villanes PantojaNo ratings yet
- Tema 7 Envolturas CelularesDocument16 pagesTema 7 Envolturas CelularesvictoriaNo ratings yet
- MEMBRANA PLASMATICADocument5 pagesMEMBRANA PLASMATICApimenteltamara2No ratings yet
- Membranas biológicas: estructura y funciones de la bicapa lipídicaDocument26 pagesMembranas biológicas: estructura y funciones de la bicapa lipídicapaolo bryan velasquez alvarezNo ratings yet
- Membrana Plasmatica Karp PDFDocument13 pagesMembrana Plasmatica Karp PDFJhon Mario RojasNo ratings yet
- BICT Tema 02 MembranaDocument9 pagesBICT Tema 02 MembranaJulia CasadoNo ratings yet
- Membrana celular: estructura y funciónDocument31 pagesMembrana celular: estructura y funciónlady tambraNo ratings yet
- Biología Sección 4Document13 pagesBiología Sección 4Choque Condori NoeliaNo ratings yet
- Apuntes Clases Ene Jun 2019Document121 pagesApuntes Clases Ene Jun 2019Kimberly MartinezNo ratings yet
- Biologia Lunes 05Document3 pagesBiologia Lunes 05brithany8523No ratings yet
- Membrana CelularDocument6 pagesMembrana Celularingrid gonzalez arrietaNo ratings yet
- Membrana Plasmatica 2020-I Mra - S6Document77 pagesMembrana Plasmatica 2020-I Mra - S6Dino P MoralesNo ratings yet
- Membrana PlasmáticaDocument20 pagesMembrana PlasmáticaJr EscobarNo ratings yet
- Unidad IV. MembranaDocument57 pagesUnidad IV. Membranamariangeles292003No ratings yet
- TEMA 1. Membrnas. - BC1Document8 pagesTEMA 1. Membrnas. - BC1juanma Giraldez MachucaNo ratings yet
- GuíamembranadesarrolladaDocument20 pagesGuíamembranadesarrolladaSamyd S BustosNo ratings yet
- Tema 3Document9 pagesTema 3Claudia Vazquez GarciaNo ratings yet
- MembranaDocument35 pagesMembranaIsa GarciaNo ratings yet
- Resumen para Estudio Membrana Plasmática. Dra - RodriguezDocument8 pagesResumen para Estudio Membrana Plasmática. Dra - RodriguezEd Carreño GarciaNo ratings yet
- 3 Membrana PlasmaticaDocument30 pages3 Membrana PlasmaticaSebastian EsnarriagaNo ratings yet
- Borrador Tipos de MembranasDocument5 pagesBorrador Tipos de MembranasJosé HuenulefNo ratings yet
- Membrana PlasmaticaDocument18 pagesMembrana PlasmaticaBorsalinoNo ratings yet
- Seminario 1Document17 pagesSeminario 1Luz Valderrama DurandNo ratings yet
- Tema 2 MEMBRANASDocument4 pagesTema 2 MEMBRANASSandra Sepúlveda TorresNo ratings yet
- Apuntes Biologia SuerteDocument3 pagesApuntes Biologia SuerteCarolina Alexandra ZEBALLOS LIPANo ratings yet
- Membrana celularDocument8 pagesMembrana celularSolange De La Hoz FlorezNo ratings yet
- Membrana celular: la barrera semipermeableDocument3 pagesMembrana celular: la barrera semipermeableMiriam Monica Silva FernandezNo ratings yet
- Membrana PlasmaticaDocument35 pagesMembrana PlasmaticafredNo ratings yet
- Superficie Celular - Membrana Plasmática - Transportes MolecularesDocument25 pagesSuperficie Celular - Membrana Plasmática - Transportes MolecularesDai DinamarcaNo ratings yet
- Captura de Pantalla 2023-10-08 A La(s) 4.52.33 P.M.Document31 pagesCaptura de Pantalla 2023-10-08 A La(s) 4.52.33 P.M.9whpc9jwvxNo ratings yet
- Clase 2 Membrana BiologiaDocument63 pagesClase 2 Membrana BiologiaSebastian PedrerosNo ratings yet
- TEMA 5. MembranasDocument6 pagesTEMA 5. MembranasIris RoigNo ratings yet
- Membrana CelularDocument16 pagesMembrana Celularfernanda salinasNo ratings yet
- Membrana celular: lípidos y proteínasDocument136 pagesMembrana celular: lípidos y proteínasRocío Bravo AcedoNo ratings yet
- Estructura de La Membrana Plasmática y Mecanismos de TransporteDocument18 pagesEstructura de La Membrana Plasmática y Mecanismos de TransporteDiego Lopez ClevesNo ratings yet
- 1 BIOMEMBRANASDocument30 pages1 BIOMEMBRANASaaaael00004910No ratings yet
- Estructura y Transporte de MembranaDocument94 pagesEstructura y Transporte de MembranaMacarena FranchiNo ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012From EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012No ratings yet
- IRDocument11 pagesIRGustavo Torres DíazNo ratings yet
- 1 Impacto Del Diseño de Plantas Biotecnológicas en La SociedadDocument5 pages1 Impacto Del Diseño de Plantas Biotecnológicas en La SociedadCarlos Salas SalgadoNo ratings yet
- 04 Laboratorio 1 Estatica 1Document15 pages04 Laboratorio 1 Estatica 1DiegoNo ratings yet
- 10 Laboratorio 7 Termodinámica - Gases IdealesDocument7 pages10 Laboratorio 7 Termodinámica - Gases IdealesGustavo Torres DíazNo ratings yet
- Unicación Efectiva PDFDocument12 pagesUnicación Efectiva PDFGustavo Torres DíazNo ratings yet
- Trabajo Grupal Semana 3Document3 pagesTrabajo Grupal Semana 3Gustavo Torres DíazNo ratings yet
- Ba 12Document8 pagesBa 12Gustavo Torres DíazNo ratings yet
- 11 Laboratorio 8 Refraccion de La Luz LentesDocument18 pages11 Laboratorio 8 Refraccion de La Luz LentesGustavo Torres DíazNo ratings yet
- Refracción de la luz en lentes y aguaDocument19 pagesRefracción de la luz en lentes y aguaGustavo Torres Díaz100% (2)
- 2 ArqDocument2 pages2 ArqGustavo Torres DíazNo ratings yet
- Práctica de Laboratorio #06 Calor Específico de SólidosDocument8 pagesPráctica de Laboratorio #06 Calor Específico de SólidosGustavo Torres DíazNo ratings yet
- Cuestionario Previo Laboratorio 5Document3 pagesCuestionario Previo Laboratorio 5Gustavo Torres DíazNo ratings yet
- 10 Laboratorio 7 Termodinámica Gases IdealesDocument13 pages10 Laboratorio 7 Termodinámica Gases IdealesGustavo Torres DíazNo ratings yet
- Mapa Mental de Partes de Una RedDocument1 pageMapa Mental de Partes de Una RedGustavo Torres DíazNo ratings yet
- Vectores de Transferencia GénicaDocument5 pagesVectores de Transferencia GénicaGustavo Torres DíazNo ratings yet
- Resultados Del Test de MemoriaDocument1 pageResultados Del Test de MemoriaGustavo Torres DíazNo ratings yet
- AbortoDocument36 pagesAbortoGustavo Torres DíazNo ratings yet
- Unicación Efectiva PDFDocument12 pagesUnicación Efectiva PDFGustavo Torres DíazNo ratings yet
- 14 Anexo 3 Manual PASCO-1Document7 pages14 Anexo 3 Manual PASCO-1Gustavo Torres DíazNo ratings yet
- Causas de la problemática del sistema penitenciario en el Perú 2000-2013Document61 pagesCausas de la problemática del sistema penitenciario en el Perú 2000-2013Gustavo Torres Díaz100% (2)
- Ideología de Género v1Document30 pagesIdeología de Género v1Gustavo Torres Díaz100% (1)
- Efecto Seebeck genera electricidadDocument4 pagesEfecto Seebeck genera electricidadGustavo Torres Díaz100% (1)
- Trip TicoDocument2 pagesTrip TicoGustavo Torres DíazNo ratings yet
- Ensayo Pil DoraDocument4 pagesEnsayo Pil DoraGustavo Torres DíazNo ratings yet
- Aromatizantes de Comidas Con Bioactividad 3Document10 pagesAromatizantes de Comidas Con Bioactividad 3Gustavo Torres DíazNo ratings yet
- A 3Document1 pageA 3Gustavo Torres DíazNo ratings yet
- La SalDocument1 pageLa SalGustavo Torres DíazNo ratings yet
- A3 - GalletasDocument1 pageA3 - GalletasGustavo Torres DíazNo ratings yet
- DX Enf InfecciosasDocument58 pagesDX Enf InfecciosasGustavo Torres DíazNo ratings yet
- Carambo LaDocument2 pagesCarambo LaGustavo Torres DíazNo ratings yet
- Guía de Práctica de Embriología General ScribdDocument12 pagesGuía de Práctica de Embriología General ScribdAnel MamaniNo ratings yet
- Desarrollo niño 0-5 añosDocument20 pagesDesarrollo niño 0-5 añosMoisés Saraya SalasNo ratings yet
- Citoesqueleto JAGTDocument5 pagesCitoesqueleto JAGTJaime GarcíaNo ratings yet
- Respuestas Del CuadernilloDocument9 pagesRespuestas Del CuadernilloJOSE DAVID CARVAJAL SICACHANo ratings yet
- Niveles organización seres vivos célula tejido órganoDocument85 pagesNiveles organización seres vivos célula tejido órganoErna Pilar GonzalezNo ratings yet
- Valor Nutritivo de Las PasturasDocument19 pagesValor Nutritivo de Las PasturasEMMANUEL ALEXANDER SESSAREGO DÁVILANo ratings yet
- Teoria QuimiosinteticaDocument2 pagesTeoria QuimiosinteticaPamela GrNo ratings yet
- Mariposa organillo claraDocument4 pagesMariposa organillo claraSebastian ArdilaNo ratings yet
- 1semiologia de La Fiebre 2020Document23 pages1semiologia de La Fiebre 2020Doctora CangrejaNo ratings yet
- ChloeDocument10 pagesChloeSovietWantanNo ratings yet
- BROCHURE ESTÉTICO Ceeluu DMEDICAL - CompressedDocument38 pagesBROCHURE ESTÉTICO Ceeluu DMEDICAL - CompressedGaro AlejandroNo ratings yet
- NeuroC 2Document18 pagesNeuroC 2Edwin ChavezNo ratings yet
- ¿Qué Es El ADN?: Características: Molécula de ADNDocument2 pages¿Qué Es El ADN?: Características: Molécula de ADNHector IrizarryNo ratings yet
- Examen Segunda Unidad BioquimicaDocument10 pagesExamen Segunda Unidad BioquimicaUrider ValladolidNo ratings yet
- Guia Acidos Nucleicos 11°Document6 pagesGuia Acidos Nucleicos 11°Doris María Correa MartínezNo ratings yet
- Clasificación de Cultivos BacterianosDocument4 pagesClasificación de Cultivos BacterianosAndres Alejandro Yela BastidasNo ratings yet
- Cadenas tróficas ecosistemasDocument5 pagesCadenas tróficas ecosistemasAnthonnythNo ratings yet
- Mejoramiento Genético de Cucurbita Moschata PDFDocument107 pagesMejoramiento Genético de Cucurbita Moschata PDFXiomara Holguín VillajuanNo ratings yet
- Estructura y composición florística de Cinchona officinalis L. en LojaDocument144 pagesEstructura y composición florística de Cinchona officinalis L. en LojaJose MorenoNo ratings yet
- Factores Del Poder ToxigenicoDocument39 pagesFactores Del Poder Toxigenicokareneliz22No ratings yet
- Guia de LaboratorioDocument24 pagesGuia de LaboratorioANDRES CAMILO TIQUE LARANo ratings yet
- Actividad Unidad 5. Pruebas para Cocos Gram Positivos.Document5 pagesActividad Unidad 5. Pruebas para Cocos Gram Positivos.ALEJANDRA BUENROSTRO AMADORNo ratings yet
- V de Gowin Extracción de ADNDocument2 pagesV de Gowin Extracción de ADNJuan Pablo Ibañez Correa71% (7)
- Tesis Sobre Historia de La EcologíaDocument108 pagesTesis Sobre Historia de La EcologíajoshBenNo ratings yet
- Parasitismo PluricelularDocument8 pagesParasitismo PluricelularMauricio Alejandro Quezada BustilloNo ratings yet
- Diapositivas de BiofarmaciaDocument33 pagesDiapositivas de Biofarmaciaroberth panchanaNo ratings yet
- Clase 2 de Biologia de 11Document7 pagesClase 2 de Biologia de 11jesus pabonNo ratings yet
- Leishmanias PDFDocument22 pagesLeishmanias PDFRoger CHNo ratings yet
- 621Document368 pages621edmundo vegaNo ratings yet
- INFECTOLOGIADocument335 pagesINFECTOLOGIAJorge Nuñez YongNo ratings yet