You might also like
- INFORME de Carbohidratos y ProteinasDocument11 pagesINFORME de Carbohidratos y ProteinasYane Rojas ZevallosNo ratings yet
- Bioquimica 1. Analisis Cuantitativo de EnzimasDocument12 pagesBioquimica 1. Analisis Cuantitativo de EnzimasangelaNo ratings yet
- Retículo Endoplasmatico LisoDocument12 pagesRetículo Endoplasmatico LisoSerbulo OLAYA CABRERA0% (1)
- Clasificacion de Productos Naturales Basado en La BiogenesisDocument3 pagesClasificacion de Productos Naturales Basado en La BiogenesisJosué AduvireNo ratings yet
- FOSFOLIPIDOSDocument6 pagesFOSFOLIPIDOSalis castilloNo ratings yet
- Célula VegetalDocument6 pagesCélula VegetalCamila Traslaviña100% (1)
- Acidos NucleicosDocument37 pagesAcidos NucleicosBrenda YepesNo ratings yet
- Terpenoides y Esteroides - 2017Document40 pagesTerpenoides y Esteroides - 2017carlos javier100% (1)
- Prueba Del IndolDocument4 pagesPrueba Del IndolEddy Castellon RodriguezNo ratings yet
- Introducción Extracción Del ADNDocument2 pagesIntroducción Extracción Del ADNAlejandra TKTNo ratings yet
- Practica 1 EnzimasDocument15 pagesPractica 1 EnzimasSamantha0523No ratings yet
- Ácidos nucleicos: estructura y funcionesDocument20 pagesÁcidos nucleicos: estructura y funcionesSherly Yesenia Melgarejo CastañedaNo ratings yet
- Proteinas y EnzimasDocument48 pagesProteinas y EnzimasWillSmith SanchezNo ratings yet
- Reconocimiento de proteínasDocument9 pagesReconocimiento de proteínasAshly FloresNo ratings yet
- CerebrosidosDocument2 pagesCerebrosidosJose Gregorio Davila HernandezNo ratings yet
- VACUOLASDocument18 pagesVACUOLASLuis Garcia Toraño100% (1)
- Extraccion Casera de AdnDocument2 pagesExtraccion Casera de AdnKatia Aramburu SoteloNo ratings yet
- Bioelementos y Biomoléculas Inorgánicas U OrgánicasDocument19 pagesBioelementos y Biomoléculas Inorgánicas U OrgánicasLEYDI ARIANA CRUZ GARCIANo ratings yet
- Extraccion e Identificacion de Lipidos PDFDocument13 pagesExtraccion e Identificacion de Lipidos PDFSofia SantanaNo ratings yet
- IntroducciónDocument5 pagesIntroducciónFabián AndrésNo ratings yet
- MUCOPOLISACARIDOSDocument3 pagesMUCOPOLISACARIDOSVictor Rafael Pachamango ChunqueNo ratings yet
- OligosacáridosDocument1 pageOligosacáridosGrupoDeApoyoIngenieriaCivilNo ratings yet
- Nº 3 Hidrólisis de PolisacáridosDocument7 pagesNº 3 Hidrólisis de PolisacáridosNicole Sanchez FigueroaNo ratings yet
- EnzimasDocument8 pagesEnzimasMARÍA VEGANo ratings yet
- Tejidos VegetalesDocument63 pagesTejidos VegetalesGary BrianNo ratings yet
- Claisen CondensaciónDocument21 pagesClaisen CondensaciónCamilo PérezNo ratings yet
- Resumen Las ProteinasDocument2 pagesResumen Las ProteinasNicolas Patricio Sitja50% (2)
- Informe de Laboratorio: Reconocimiento de BiomoléculasDocument24 pagesInforme de Laboratorio: Reconocimiento de BiomoléculasAndresNo ratings yet
- BasesNitrogenadas PDFDocument47 pagesBasesNitrogenadas PDFAlfonso Martínez BravoNo ratings yet
- Práctica 4 - Identificación de Carbohidratos - G7Document11 pagesPráctica 4 - Identificación de Carbohidratos - G7Juan ParedesNo ratings yet
- Resumen - Biologia ADN y ARNDocument2 pagesResumen - Biologia ADN y ARNDaniela ChirinosNo ratings yet
- Fracionamiento Celular LaboratorioDocument13 pagesFracionamiento Celular LaboratorioYeisyAlayoNo ratings yet
- Practica 1. Observación e Identificación de ProtozoariosDocument9 pagesPractica 1. Observación e Identificación de ProtozoariosDianis Rangel0% (1)
- Biotecnología RojaDocument3 pagesBiotecnología RojaCarlos SerranoNo ratings yet
- MACROELEMENTOSDocument10 pagesMACROELEMENTOSVictor SalazarNo ratings yet
- Aislamiento Del Adn!!Document6 pagesAislamiento Del Adn!!Ferc RiversNo ratings yet
- Guia 5 - Permeabilidad Celular - ERITROCITOS Y ELODEADocument8 pagesGuia 5 - Permeabilidad Celular - ERITROCITOS Y ELODEANIKOLE SANABRIA RODRIGUEZNo ratings yet
- Taller de ProteínasDocument9 pagesTaller de ProteínasVALENTINA ANDRADE ANDRADE100% (1)
- Inclusiones CitoplasmaticasDocument5 pagesInclusiones CitoplasmaticasAngie Jugo TorresNo ratings yet
- Tejidos VegetalesDocument51 pagesTejidos VegetalesStefy Recalde80% (5)
- Marco Teorico BiomoleculasDocument4 pagesMarco Teorico BiomoleculasCarlos Francisco Alvarado RodriguezNo ratings yet
- CarbohidratosDocument6 pagesCarbohidratosMontserrat VergaraNo ratings yet
- 15 Preguntas LipidosDocument5 pages15 Preguntas LipidosKassandra VegaNo ratings yet
- Identificación carbohidratosDocument26 pagesIdentificación carbohidratosAmada100% (1)
- Practica 2 Carbohidratos y LipidosDocument7 pagesPractica 2 Carbohidratos y Lipidosmelissa turriagoNo ratings yet
- Citoesqueleto ForoDocument39 pagesCitoesqueleto ForoClaudia Montes GuerraNo ratings yet
- LipidosDocument22 pagesLipidosRotcehLovegoodNo ratings yet
- Cuesionario RecomendacionesDocument4 pagesCuesionario RecomendacionesSantiagoCulquiNo ratings yet
- Informe Lab Citología MohosDocument6 pagesInforme Lab Citología MohosFernando VillegasNo ratings yet
- HISTIDINADocument9 pagesHISTIDINAJosefina MateoNo ratings yet
- Inclusiones Citoplasmaticas PDFDocument10 pagesInclusiones Citoplasmaticas PDFAndrea Casasnovas50% (2)
- Esteroles (Colesterol)Document8 pagesEsteroles (Colesterol)Brando Molina NuñezNo ratings yet
- Historia de La Bioquímica y Su ImportanciaDocument7 pagesHistoria de La Bioquímica y Su ImportanciageynerNo ratings yet
- Metabolismo (Generalidades)Document5 pagesMetabolismo (Generalidades)J Andres WolfNo ratings yet
- GLUCONEOGÉNESISDocument14 pagesGLUCONEOGÉNESISnamames jodeteNo ratings yet
- Bioelementos y BiomoléculasDocument3 pagesBioelementos y BiomoléculasJavi García Fdez-MedinaNo ratings yet
- Ácidos NucleicosDocument14 pagesÁcidos Nucleicosthyago pereyraNo ratings yet
- Lectura Acidos Nucleicos 2022Document9 pagesLectura Acidos Nucleicos 2022Naomy Burgos GonzalezNo ratings yet
- GuiaDocument13 pagesGuiaMichiNo ratings yet
- Resumen Final Biol. Cel. y Molec.Document211 pagesResumen Final Biol. Cel. y Molec.Hugo MedusNo ratings yet
- ENOVA 6°2° PresentaciónDocument9 pagesENOVA 6°2° PresentaciónPatricia BusteloNo ratings yet
- F09. Tiro Vertical y Caída LibreDocument3 pagesF09. Tiro Vertical y Caída LibrePatricia BusteloNo ratings yet
- Modelos AtómicosDocument21 pagesModelos AtómicosPatricia BusteloNo ratings yet
- Capítulo 7 - Iones ComplejosDocument25 pagesCapítulo 7 - Iones ComplejosPatricia Bustelo100% (1)
- Composicion Quimica Del PlanetaDocument66 pagesComposicion Quimica Del PlanetaPatricia BusteloNo ratings yet
- Anexo 1 Teoría Atómico-MolecularDocument8 pagesAnexo 1 Teoría Atómico-MolecularPatricia BusteloNo ratings yet
- Composicion Quimica Del PlanetaDocument66 pagesComposicion Quimica Del PlanetaPatricia BusteloNo ratings yet
- Documento de 80 líneasDocument5 pagesDocumento de 80 líneasPatricia BusteloNo ratings yet
- Cap 15 Metabolismo de AminoácidosDocument45 pagesCap 15 Metabolismo de AminoácidosPatricia BusteloNo ratings yet
- Maquinas SimplesDocument53 pagesMaquinas SimplesPatricia BusteloNo ratings yet
- Composicion Quimica Del PlanetaDocument66 pagesComposicion Quimica Del PlanetaPatricia BusteloNo ratings yet
- Capítulo 1 - Química Del CarbonoDocument8 pagesCapítulo 1 - Química Del CarbonoPatricia BusteloNo ratings yet
- Biofísica 2018Document31 pagesBiofísica 2018Patricia BusteloNo ratings yet
- Propiedades ColigativasDocument20 pagesPropiedades ColigativasPatricia BusteloNo ratings yet
- Krahg Helge Generaciones Cuanticas PDFDocument455 pagesKrahg Helge Generaciones Cuanticas PDFPatricia Bustelo100% (1)
- Biofísica 2018Document31 pagesBiofísica 2018Patricia BusteloNo ratings yet
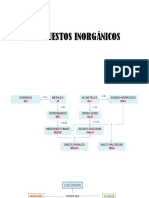
- Fórmula y Nomenclatura Compuestos InorgánicosDocument9 pagesFórmula y Nomenclatura Compuestos InorgánicosPatricia BusteloNo ratings yet
- Biofísica 2018Document2 pagesBiofísica 2018Patricia BusteloNo ratings yet
- RespuestasDocument82 pagesRespuestasPatricia BusteloNo ratings yet
- Geografía AmbientalDocument51 pagesGeografía AmbientalPatricia BusteloNo ratings yet
- Biofísica 2018Document31 pagesBiofísica 2018Patricia BusteloNo ratings yet
- Aplicación de Ondas en MedicinaDocument15 pagesAplicación de Ondas en MedicinaPatricia BusteloNo ratings yet
- PLAN de CLASE (Para Completar)Document3 pagesPLAN de CLASE (Para Completar)Patricia BusteloNo ratings yet
- Compuestos InorgánicosDocument7 pagesCompuestos InorgánicosPatricia BusteloNo ratings yet
- Compuestos InorgánicosDocument10 pagesCompuestos InorgánicosPatricia BusteloNo ratings yet
- La Corrosión Del AluminioDocument3 pagesLa Corrosión Del AluminioPatricia BusteloNo ratings yet
- Propiedades ColigativasDocument1 pagePropiedades ColigativasCarlosAndresBuelvasRodriguezNo ratings yet
- Tipos de Reacciones QuímicasDocument26 pagesTipos de Reacciones QuímicasPatricia BusteloNo ratings yet
- TP N°-Propiedades PeriódicasDocument2 pagesTP N°-Propiedades PeriódicasPatricia BusteloNo ratings yet
- Nivel SubmicroscópicoDocument20 pagesNivel SubmicroscópicoPatricia Bustelo100% (1)
- Curso Valvulas de ControlDocument90 pagesCurso Valvulas de ControlJuana Lucia Cruz Arias100% (1)
- EulogenolDocument3 pagesEulogenollaura segundoNo ratings yet
- INFORME N - 3 Uso y Majeno Del Banco H.Document31 pagesINFORME N - 3 Uso y Majeno Del Banco H.Manuel SerranoNo ratings yet
- Estructuras Complementarias en CanalesDocument10 pagesEstructuras Complementarias en CanalesJerson BenitesNo ratings yet
- Práctico GaruttiDocument6 pagesPráctico GaruttiAgustinhillmanNo ratings yet
- 02 AnalisisMateriaEnergia AMAE-03 Rev PDFDocument28 pages02 AnalisisMateriaEnergia AMAE-03 Rev PDFLuispanqueNo ratings yet
- Central CarboeléctricaDocument10 pagesCentral Carboeléctricabeto117kNo ratings yet
- Practica Cristalizacion 2015Document14 pagesPractica Cristalizacion 2015Reynel Giron GomezNo ratings yet
- 1 Proteobacterias Bacterias Fototrofas PDFDocument13 pages1 Proteobacterias Bacterias Fototrofas PDFrossi maussaNo ratings yet
- Desencolado (02.04Document64 pagesDesencolado (02.04elvis bellidoNo ratings yet
- Respiración celular crucigramaDocument1 pageRespiración celular crucigramaMauricio Ivan Sanchez88% (8)
- Preparación de soluciones acuosas optimizadas paraDocument3 pagesPreparación de soluciones acuosas optimizadas paraNICOLAS JARAMILLO MAYORNo ratings yet
- Preparac LiquidasDocument40 pagesPreparac LiquidasAugusto Tamani0% (1)
- AcetilenoDocument11 pagesAcetilenoYameris MoralesNo ratings yet
- 8.1 ExtraccionDocument17 pages8.1 ExtraccionAnonymous ZaxvppAHNo ratings yet
- Evaluaciones CSIDocument17 pagesEvaluaciones CSIAlex GarciaNo ratings yet
- Electrodo rutiloDocument6 pagesElectrodo rutiloDarnely RivasNo ratings yet
- Cedva 1Document20 pagesCedva 1Rafael Sosa CanoNo ratings yet
- BombasDocument11 pagesBombasraul cabanillasNo ratings yet
- 05 Salinidad en Suelos EspanolDocument11 pages05 Salinidad en Suelos EspanolKarín Díaz CarriónNo ratings yet
- Aerenquima y ParenquimaDocument8 pagesAerenquima y ParenquimaJose Antonio Prado Martines100% (1)
- Aplicación Del Método Del Hidrograma Unitario TriangularDocument2 pagesAplicación Del Método Del Hidrograma Unitario TriangularFidelmar MerlosNo ratings yet
- Espectro MasasDocument36 pagesEspectro MasasMarco Dueñas FloresNo ratings yet
- Electro Bomb AsDocument9 pagesElectro Bomb AsMarii AcostaNo ratings yet
- Especificaciones Tecnicas Veredas CacazuDocument34 pagesEspecificaciones Tecnicas Veredas CacazuJeffsrcNo ratings yet
- Soluciones químicas: propiedades y tiposDocument5 pagesSoluciones químicas: propiedades y tiposIsaac Rodriguez100% (1)
- Fluidos de PerforacionDocument57 pagesFluidos de PerforacionWilmer SuarezNo ratings yet
- MBRACE FolletoDocument8 pagesMBRACE FolletoJorge Infantas MotaNo ratings yet